

Старение — рост риска смерти от естественных причин с течением времени, биологический процесс постепенного нарушения и потери важных функций организма или его частей, в частности способности к размножению и регенерации. Вследствие старения организм становится менее приспособленным к условиям окружающей среды, уменьшает и теряет свою способность бороться с хищниками и противостоять болезням и травмам. Наука, которая изучает старение человека, называется геронтологией, а её раздел, который изучает непосредственно биологическую сторону старения, носит название биогеронтология.
Эволюция старения является одной из нерешённых загадок биологии. Существует много теорий, объясняющих старение с точки зрения эволюции. В общем случае считается, что главной движущей силой скорости старения является экзогенная смертность в условиях сурового окружающего мира, из-за которой эволюционное давление практически не способствует отбору изменений, замедляющих старение. Согласно теории накопления мутаций вредные мутации, которые оказывают негативные эффекты в позднем возрасте (после репродуктивного периода), могут пассивно накапливаться без какого-либо сопротивления со стороны естественного отбора. Теория антагонистической плейотропии подходит к вопросу с другой стороны, предполагая, что если старение станет редким явлением, то естественный отбор будет идти в пользу генов, которые дают больше преимуществ в начале жизни, но могут оказывать негативный эффект в позднем возрасте. Объединяет эти теории то, что повышение экзогенной смертности приводит к эволюционному сокращению продолжительности жизни, и наоборот. Однако эти теории не универсальные, и бывают исключения, а наличие экзогенной смертности среди некоторых теоретически бессмертных существ противоречит классическим теориям.
Термин «старение» может использоваться для описания социальных эффектов старения человека, а также для описания разрушения неживых систем, например, старение металлов.
Старение разных организмов
Явление старения в той или иной мере наблюдается практически у всех живых организмов, за исключением ряда организмов с пренебрежимым старением. У людей в возрасте около 100 лет также наблюдается явление пренебрежимого старения.
Старение и размножение
Размножение — важная функция жизни организмов: все другие жизненные процессы, в частности старение и смерть, адаптированы к его потребностям. Для понимания процесса старения важно подчеркнуть различия между одноплодными и многоплодными стратегиями размножения. Одноплодные организмы размножаются с помощью единичного акта размножения на протяжении жизни организма. Большинство одноплодных организмов — однолетние и двулетние растения, но среди них много насекомых и несколько позвоночных животных, таких как розовый лосось и угорь. Многоплодные организмы, с другой стороны, размножаются периодически на протяжении диапазона половой зрелости, который обычно составляет большую часть жизни.
Одноплодные организмы
У одноплодных форм размножения старение имеет место в конце жизненного периода, после которого организм проходит через стадию очень быстрого старения, которое приводит к смерти организма. У растений фаза старения обычно является составляющей частью возобновляемого процесса, важной для его завершения. Распространение семян, например, сопровождается процессами созревания, падения плодов и высыхания семян — процессов, неотделимых от процесса старения растения. Это быстрое старение вызывается с помощью определённых гормонов, уровень которых меняется в зависимости от состояния развития растения и экологических условий. Если, например, препятствовать выделению гормона роста растений — ауксина — экспериментальными методами, растение живёт дольше и проходит через нетипичное для него долгое старение.
Подобная картина старения наблюдается и у некоторых насекомых, которые отличаются двумя чёткими стратегиями адаптивной окраски: защитная окраска, с помощью которой насекомые маскируются от потенциальных хищников, и отпугивающая окраска, которая обычно указывает на ядовитость насекомого. Эти два примера адаптации имеют разные оптимальные стратегии выживания видов: насекомые с защитной окраской умирают быстрее после завершения размножения, тем самым сокращая возможность для хищников научиться распознавать их,— тогда как насекомые с отпугивающей окраской живут очень долго после размножения, увеличивая возможность хищников запомнить их. Обе стратегии адаптаций найдены среди представителей семейства мотыльков-сатурний, на примере которых было показано, что продолжительность их жизни после размножения контролируется гормональной системой, которая также контролирует ту часть времени, которую насекомые проводят в полёте: мотыльки с защитной окраской проводят больше времени в полёте, истощают себя и быстро умирают, тогда как мотыльки с отпугивающей окраской летают меньше, сохраняют энергию, и в результате — живут дольше.
Многоплодные организмы
Многоплодные формы включают большинство позвоночных животных, большинство долгоживущих насекомых, ракообразных и пауков, головоногих и брюхоногих моллюсков и многолетние растения. В отличие от одноплодных форм, многоплодным организмам не нужно использовать все жизненные силы до конца их воспроизводящей фазы (фазы размножения) для того, чтобы размножение прошло успешно, и средняя продолжительность жизни относительно периода размножения весьма значительно меняется у разных особей и в зависимости от вида: маленькие грызуны и дикие птицы используют в среднем только от 10 до 20 процентов их потенциального периода размножения, тогда как киты, слоны, обезьяны и другие крупные млекопитающие в природных условиях используют более 50 процентов своих периодов размножения, и часто даже переживают его.
У многоплодных организмов старение имеет постепенный характер: без очевидных специфических системных или экологических механизмов, которые бы инициировали этот процесс. Старение в первую очередь проявляется как снижение возможностей организма к размножению. У видов, которые достигают фиксированного размера тела, снижение возможностей к размножению начинается довольно рано — и ускоряется с возрастом. У больших пресмыкающихся, которые достигают половой зрелости ещё имея небольшой размер и продолжают расти на протяжении долгого воспроизводящего периода, число откладываемых яиц увеличивается с возрастом на протяжении довольно долгого периода, но в конце концов прекращает расти и постепенно снижается. Воспроизводящий период в таких случаях короче продолжительности жизни.
Ещё одной особенностью многоплодных организмов является зависимость скорости процесса старения от способности размножаться и от возможностей выживания потомства — наиболее плодовитые организмы стареют намного быстрее, чем менее плодовитые. Такая зависимость позволяет выживать животным с малой продолжительностью жизни за счёт большого потомства, а неплодовитым животным — за счёт большого числа возможностей для размножения.
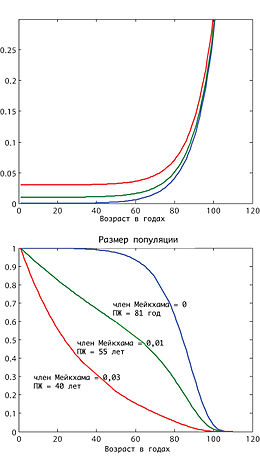
Для математического описания старения многоплодных организмов иногда используется так называемый закон смертности Гомпертца — Мейкхама (или просто Гомпертца), согласно которому вероятность смерти экспоненциально возрастает с возрастом: , где x — возраст, а p — относительная вероятность смерти за определённый промежуток времени, a и b — коэффициенты. Таким образом, при отсутствии постоянного члена a, размер популяции уменьшается с возрастом по двойной экспоненте . Этот закон эмпирический и имеет место не для всех животных и не на всех промежутках времени, но он наиболее простой для сравнения старения разных организмов, и потому коэффициент b часто используется в качестве показателя скорости старения.

![s(x)=exp[-m(b^{x}-1)]](https://wikimedia.org/api/rest_v1/media/math/render/svg/c26ed314af895c5a482ff82d45685acbacc6a108)
Закон Гомпертца — Мейкхама является только приближением, верным в среднем возрастном диапазоне. В области малого возраста наблюдается значительно более высокая смертность, чем предусматривается этим законом. Например, северная треска во время нереста может откладывать до 6 миллионов икринок, но только небольшое количество из них выживает до момента половой зрелости. Такая смертность является преимущественно результатом неспособности молодых особей избегать хищников, бороться с болезнями, а также может быть результатом врождённых дефектов, и не является результатом старения. В области позднего возраста, наоборот, наблюдается уменьшения смертности по сравнению с законом Гомпертца, точнее выход вероятности смерти за единицу времени на плато. Как и в случае детской смертности, это общий закон, который наблюдается даже в неживой природе. И хотя одним из возможных объяснений этого явления могла бы быть гетерогенность популяции, современные данные чётко указывают на связь выхода смертности на плато с замедлением процесса старения.
Для многоплодных организмов очевидно влияние факторов популяционной динамики на эволюцию репликативного (клеток генеративной линии) и соматичного (телесного) старения. Пропорциональный вклад индивидуума в скорость увеличения размера многоплодной популяции уменьшается с возрастом по мере уменьшения способности размножаться. Эти факты предполагают наличие оптимального размера потомства от одной особи. Насколько это может направить эволюцию специфического старения или методов защиты против старения, вызванного другими причинами, — всё ещё остаётся открытым вопросом, в котором нет единого мнения у исследователей.
Старение в разных таксономических группах
Многоклеточные животные
Эффект старения животных обычно наблюдается только в защищённых средах (например, лабораториях и зоопарках). В природных условиях доминирующей причиной смерти являются хищники, болезни или нехватка еды, поэтому старение проявляется очень редко. Как правило, естественная продолжительность жизни животного только ненамного превышает время достижения половой зрелости, а максимальная продолжительность жизни до смерти от старости в несколько раз больше (однако, существует несколько исключений из этого правила). В следующей таблице приведены значения максимальной зарегистрированной продолжительности жизни некоторых организмов до смерти от старости, таблица составлена преимущественно основываясь на данных, полученных в защищённых условиях.
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
- Примечания:
- а Эта таблица является сокращённым вариантом таблицы, приведенной в статье Максимальная продолжительность жизни, с целью показать приблизительный диапазон продолжительностей жизни для основных групп организмов. Ссылки на источники каждого значения приведены в полном варианте. При составлении таблицы указывались все найденные значения возраста, хотя достоверность их разная. Лучшим источником возраста для большинства организмов является AnAge Database в связи с тем, что эта база данных в большинстве случаев содержит подробные ссылки на первоисточники использованной информации и оценивает их достоверность.
.
- б Человек способен пользоваться услугами медицины, кроме того, случаи долгой жизни человека намного лучше задокументированы. В результате продолжительность жизни человека кажется дольше, чем у аналогичных животных.
- с личинка подёнки живёт до 2—3 лет.
- Факторы, которые оказывают влияние на продолжительность жизни.
Продолжительность жизни очень сильно разнится между видами животных. Несколько факторов имеют своё влияние на неё. В большинстве случаев продолжительность жизни зависит от плодовитости животного: чем больше потомства оно даёт, тем меньше живёт. Кроме того, продолжительность жизни зависит от размера животного, размера его мозга и метаболической активности. Например, как правило меньшие по размеру животные имеют меньшую, а большие по размеру — большую продолжительность жизни.
Простейшие животные
Некоторые низшие животные практически избегают старения за счёт быстрого обновления клеток всех тканей своего тела, что возможно за счёт простой и децентрализованной структуры тела. Примерами таких организмов являются морские актинии и пресноводные гидры. В исследовании, опубликованном в 1998 году, было показано, что по внешним, цитологическим признакам и способности к размножению, гидра не проявляет признаков старения на протяжении всего исследования в течение 4 лет. Даже если она, в принципе, и может стареть, разница между продолжительностью жизни и временем достижения половой зрелости, который составляет лишь около недели, весьма существенна.
Медленно стареющие позвоночные животные
Хотя у большинства животных старение очевидно, у некоторых животных оно незначительно. Примерами таких животных являются некоторые крупные пресмыкающиеся, в первую очередь черепахи. Например, галапагосская черепаха (Geochelone nigra) способна жить до 177 лет, а некоторые рыбы, например осетровые, живут дольше, чем 150 лет. Тем не менее, продолжительность жизни и старение этих животных исследованы очень плохо. Кроме того, даже на протяжении долгой жизни, затраты энергии этих организмов незначительны, намного меньшие, чем у млекопитающих. Кроме того, эти животные постепенно увеличиваются в размере даже после достижения половой зрелости, что позволяет им разбавлять стареющие клетки тела новыми.
Млекопитающие
Лучше всего исследована зависимость продолжительности жизни от таксономической группы среди млекопитающих. Приматы, в целом, — наиболее долгоживущая группа, хотя у некоторых небольших обезьян Нового Света короткая продолжительность жизни. Мышиные грызуны недолговечны, тогда как беличьи грызуны достигают втрое большего возраста, чем мышиные. Продолжительность жизни зависит от трёх отдельных факторов: веса мозга, веса тела, и скорости обмена веществ (метаболизма). Зависимость продолжительности жизни от этих факторов может быть выражена в форме эмпирического уравнения: L = 5,5 E0,54 S−0,34 M−0,42. Продолжительность жизни (L) млекопитающих в годах зависит от массы мозга (E) и массы тела (S) в граммах и от скорости обмена веществ (М) в калориях на грамм за час. Позитивный показатель степени для E (0,54) показывает, что долговечность млекопитающих сильно и положительно коррелирует с размером мозга независимо от размера тела или скорости обмена веществ. Отрицательный коэффициент корреляции со скоростью обмена веществ показывает, что продолжительность жизни уменьшается с ростом активности организма. Отрицательная корреляция с весом тела указывает, что тенденция, что большие животные живут дольше, является результатом не большого размера тела, а большей массы мозга.
Типичная зависимость несколько нарушается в случае пород собак. Большие по размеру породы собак, хотя и достигают половой зрелости медленнее, живут значительно меньше, разница составляет около 2 раз между крупнейшими и самыми мелкими породами. Этот пример был первым наблюдением, которое показало, что селекция в принципе способна изменять продолжительность жизни.
Птицы
Такой же вид зависимости L от E, S и М справедлив и для птиц, но птицы в целом живут дольше, чем млекопитающие, несмотря на более высокие температуры тела и скорости метаболизма.
Пресмыкающиеся
Продолжительности жизни крупных пресмыкающихся превышает продолжительности жизни млекопитающих соответствующего размера, но скорость их естественного обмена веществ в десятки раз меньше. Таким образом, полная потеря энергии за жизнь у пресмыкающихся меньше, чем у млекопитающих.
Членистоногие
Продолжительность жизни разных видов членистоногих может составлять от нескольких дней до нескольких десятилетий. Наиболее недолгоживущие насекомые имеют одну короткую воспроизводящую фазу; долгоживущие пауки и ракообразные — многоплодные, с годовыми циклами воспроизводства.
Затраты энергии
Крупные животные с развитым мозгом, особенно приматы, вырабатывают на протяжении жизни наибольшее количество энергии. Например, полная выработка энергии за жизнь человека на грамм ткани составляет около 1,2 млн калорий, а за жизнь таких домашних животных, как кошки и собаки, только 0,4 млн калорий.
Приведённая выше зависимость справедлива для теплокровных животных. Хладнокровные животные, которые впадают в периоды ежедневной неактивности или зимней спячки, могут значительно сокращать свою метаболическую активность, часто более чем в десять раз. Насекомоядные летучие мыши умеренных широт — известнейший пример — хотя они живут более 20 лет, почти 80 процентов этого времени тратится в глубокой спячке. В результате потеря ими энергии на протяжении жизни не превышает затраты энергии другими маленькими млекопитающими.
Одноплодные животные
Как уже указывалось, картина старения одноплодных организмов очень отличается от млекопитающих. Одним из самых интересных примеров такого старения является жизненный цикл розового лосося. Обычная продолжительность жизни рыбы составляет около двух лет. Мальки лосося вылупляются в пресноводных водоёмах, мигрируют вниз по течению к морю, где и проводят большую часть жизни. После достижения половой зрелости, лосось большими косяками поднимается вверх по течению к местам нереста, во время этого путешествия старость рыб ещё не заметна, а рыбы должны быть в отличной физической форме, чтобы подниматься через пороги. Однако уже через неделю после нереста, лосось умирает от старости, что легко заметить по деградации почти всех тканей организма. Хотя продолжительность жизни значительно отличается у родственных видов, все они имеют только одну возможность нереста, после которой все рыбы умирают.
Аналогичную картину старения имеет и периодическая цикада (Magicicada spp.), также известная как «17-летняя саранча». Её личинки живут в земле 17 лет, после чего превращаются во взрослых крылатых насекомых, спариваются, откладывают яйца, и умирают. Обычно все насекомые в «выводке» на территории сотен тысяч км² достигают взрослого состояния в течение 24-часового периода, что очевидно является стратегией выживания, нацеленной на то, что птицы и другие хищники не смогут справиться с огромным количеством цикад, кроме того, облегчается поиск партнера для спаривания. Продолжительность жизни взрослых насекомых составляет лишь несколько недель.
Эти примеры иллюстрируют случай, когда у одноплодных форм, которым полная сила и функциональность необходимы фактически до конца жизни, старение проходит очень быстро и связано с завершением процесса размножения, и часто активно управляется относительно простыми гормональными механизмами, которые могут развиваться под действием естественного отбора. Такие специфические генетически контролируемые процессы старения — образец запрограммированного завершения жизни, которые проявляются, когда такое старение даёт преимущество для выживания молодёжи. Но в других случаях быстрое старение одноплодных организмов является результатом истощения организмов, более не нужных после размножения.
Изнашивание незаменимых органов. Для некоторых животных причиной смерти может стать изнашивание важных незаменимых органов, даже при незначительном старении остального организма. Например, продолжительность жизни некоторых травоядных животных ограничивается стачиванием их зубов во время пережёвывания жёсткой травы. В то же время человек может вырастить только два набора зубов, а наиболее долгоживущее травоядное животное, слон, может вырастить шесть, что позволяет заменять сточенные зубы новыми. Тем не менее, в связи с относительно медленным старением и небольшим количеством естественных хищников, слоны часто доживают до возраста, когда последний набор зубов стачивается, и животное умирает от голода.
Растения
| Организм | Продолжительность жизни (оценка) |
|---|---|
| Ларрея трёхзубчатая (Larrea tridentata) | 11 000* |
| Секвойядендрон гигантский (Sequoiadendron giganteum) | 4000 |
| Дуб черешчатый (Quercus robur) | 2000 |
| Лиственница европейская (Larix decidua) | 700 |
| Черемша (Allium ursinum) | 8—10 |
| Молочная вика (Astragalus utahensis) | 3 |
- Примечание: * В этом случае возможны проблемы с идентификацией того же «индивидуума», так как имеется в виду возраст плантации клонов.
В случае растений старение организма выражено не так чётко, как у животных. Такие понятия, как старение, созревание, смерть, развитие, хлороз, некроз, высушивание, плохо разграничены и часто перекрываются. Кроме того, неясно, что делать в случае семян, которые могут сохраняться довольно долго, не проявляя никакой физиологической активности, или с частями растений, которые дают начало новым растениям в результате вегетативного размножения. Продолжительность жизни очень сильно разнится среди представителей царства растений. Некоторые виды деревьев могут жить несколько сотен лет, тогда как некоторые травянистые растения проходят жизненный цикл за несколько недель.

Рассматривая продолжительность жизни растений, важно принимать во внимание, что смертность клеток очень высока на протяжении всей жизни растения, а отмирание тканей, или, в случае одноплодных организмов, целых растений, регулируется системой гормонов. Рост сосудистых растений зависит от активности меристем, которые состоят из клеток, аналогичных стволовым клеткам животных, и по сути представляют собой эмбриональные ткани. В случае многих многоплодных (многолетних) растений, этот рост может продолжаться практически неограниченно. Остальные ткани растений, тем не менее, постоянно отмирают. Например, большая часть массы дерева состоит из мёртвых клеток древесины. Отдельные органы растений, такие как листья, имеют продолжительность жизни значительно меньше, чем продолжительность жизни всего организма. Листву выгодно поддерживать, только если она способствует выживанию всего растения. Это видно на примере листопадных растений умеренного пояса: при уменьшении светового дня или снижении температуры, в растениях запускается запрограммированная смерть клеток, которая обычно приводит к смене окраски листьев и их опаданию.
Это «желтение» листьев в литературе часто называется их старением или синдромом старения. С помощью процесса старения питательные вещества умирающего листа мобилизуются для использования другими частями растения, что позволяет поддерживать их рост. Наличие питательных веществ, в первую очередь азота, является главным лимитирующим фактором роста. Кроме того, растения не могут изменять своё положение в грунте, и таким образом используют доступные минеральные вещества. В результате существует сильное эволюционное давление на развитие систем вторичной переработки необходимых для жизни компонентов.
Есть много факторов, которые могут инициировать программу старения и переработки питательных веществ. У некоторых видов её может вызвать потребность в питательных веществах в другом месте, например, для развития семян. Также программа отмирания листвы или других частей растения может быть вызвана постепенным снижением их продуктивности со временем за счёт старения фотосинтетического аппарата или сосудистой системы этих частей организма, аналогично процессу старения животных.
Процесс старения, аналогичный старению многоплодных животных, также случается у растений при отсутствии запрограммированного старения. Этот процесс характеризуется изменениями клеточной структуры, подобными изменениям структуры животных клеток. Наблюдается уменьшение пахучести для растений с продуцированием запаха.
Одноклеточные организмы
Одноклеточные организмы, как и многоклеточные, стареют. Они интенсивно исследуются из-за подобности их старения клеточному старению многоклеточных организмов. Модельным организмом для изучения клеточного старения являются почкующиеся дрожжи, (Saccharomyces cerevisiae), которые, как эукариоты, обладают биохимическими процессами, подобными биохимическим процессам стареющих клеток высших животных и растений. Также исследования проводились и среди некоторых бактерий, в частности кишечной палочки и Caulobacter crescentus.
На одноклеточных организмах были исследованы две разные формы процессов старения. Первая — старение, специфичное для материнской клетки при клеточном делении, при котором материнская клетка стареет, тогда как дочерняя клетка при каждом делении обнуляет клеточные часы (омолаживается). Старение материнской клетки проявляется в постепенном замедлении клеточного цикла, после чего клетка теряет способность к новым делениям, так называемое репликативное старение. Этот процесс был сначала найден у клеток с асимметричным делением, то есть в почкующихся дрожжах, а потом у асимметричной бактерии Caulobacter crescentus, где определение материнской клетки очевидно. Позднее наблюдения за делением клеток позволили определить материнскую клетку и в организме с симметричным делением — бактерии E. coli, где материнской клеткой является клетка со «старым концом», хотя позднее были найдены и незначительные морфологические различия между материнскими и дочерними клетками.
Возраст в данном случае определяется по числу делений, через которые прошла клетка, а не календарным временем. Типичная средняя продолжительность жизни лабораторных дрожжей (Saccharomyces cerevisiae) дикого типа составляет около 25 поколений. Функция распределения продолжительности жизни индивидуальных клеток следует закону Гомпертца, также, как и продолжительность жизни высших животных. У бактерии Caulobacter crescentus средняя продолжительность жизни составляет от 100 до 130 делений.
Второй процесс старения, также найденный в этих трёх модельных организмах, — хронологическое старение клеток, больше известный как условное старение (в связи с тем, что этот процесс проявляется только в определённых условиях окружающей среды). Этот процесс проявляется в постепенной деградации и потере жизнеспособности клеток на протяжении стационарной фазы.
На примере дрожжей было обнаружено, что, подобно процессу репликативного старения, специфичного для материнских клеток, условное старение является как процессом износа внутриклеточных структур, так и генетической программы. Например, клеточная стенка в стационарной фазе проходит через ряд генетически запрограммированных структурных изменений с целью повышения жизнеспособности и продолжительности жизни организма. Но, в конце концов клетка стареет и умирает.
Наследование долголетия
Наследование долголетия в популяциях таких животных, как плодовые мухи и мыши, может быть определено, если сравнивать таблицы продолжительности жизни естественных популяций и некоторых мутантов и гибридов. Согласно экспериментальным данным, около 30 процентов вариации долголетия самок и 20 процентов долголетия самцов определяется генетически. Эти значения подобны наследованию таких физиологических черт, как количество отложенных яиц и производство молока домашними животными.
Коэффициент экспоненты функции Гомпертца указывает скорость старения. Различия в долголетии между видами является результатом прежде всего различий в скорости старения, и потому выражаются в различиях в этом коэффициенте.
Сравнение таблиц смертности разных линий мышей одного вида показывает, что различия между линиями в первую очередь определяется различиями в независимом от возраста члене функции Гомпертца. Если линии отличаются только независимым от возраста членом, менее долголетние линии имеют большую смертность, которая выше на постоянную величину на протяжении всей жизни, что проявляется в вертикальном сдвиге функции Гомпертца. При этом часто случается, что гибриды первого поколения (F1) двух природных линий живут дольше, чем любой из родителей. Хотя исследований биохимических процессов таких гибридов не проводилось, таблицы продолжительности жизни указывают, что гибриды отличаются от родительских линий только независимым от возраста членом, но не изменением скорости старения. Другие исследования также показали, что в значительной мере вариации в продолжительности жизни между линиями мышей поясняются различиями в унаследованной склонности к определённым болезням.
Наследование долголетия людей сложнее исследовать, потому что на продолжительность жизни влияют социально-экономические и другие внешние факторы, которые усложняют корреляции между близкими родственниками. Тем не менее, исследования указывают на некоторую, хотя и небольшую, наследуемость продолжительности жизни и склонности к таким болезням как рак и болезни сердца, в первую очередь из-за того, что однояйцевые (генетически идентичные) близнецы чаще имеют схожие частоты этих заболеваний, чем соответственно разнояйцевые (генетически разные) близнецы.
Изменения организма во время старения
Физиология старения
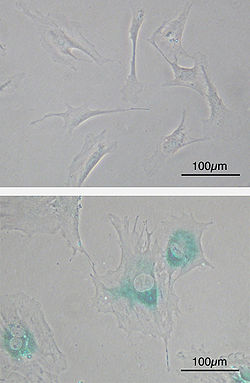
Верх: Первичные мышиные фибробласты (MEF) до старения.
Низ: MEF стареют после достижения предела Хейфлика. Клетки увеличиваются, становятся плоскими и экспрессируют новые гены, характерные для старения (SABG, синий цвет)
Наилучшим образом изучены изменения, происходящие в организмах млекопитающих, в первую очередь из-за родства этих организмов человеку, но также и в связи с тем, что симптомы старения наиболее чётко выражены у этих видов.
Все млекопитающие являются многоплодными организмами, у которых старение протекает медленно и охватывает практически все системы организма. Общие изменения включают снижение основной массы тела (живых клеток и костей) при возрастании общей массы за счёт возрастания количества жировых отложений и содержания воды. Основной обмен, то есть минимальный уровень метаболической активности, снижается во всех тканях, как и уровень добровольной активности, но последняя сильно зависит от вида животных и физической нагрузки на протяжении предыдущего периода.
Старение клеток
На клеточном уровне старение проявляется в замедлении деления клеток. Частично этот эффект является результатом так называемого предела Хейфлика деления соматических клеток. Эта граница связана с отсутствием активной теломеразы, в результате чего конечные участки хромосом, теломеры, сокращаются при каждом делении. У человека соматическая клетка может делиться около 52 раз, после чего теломеры исчезают, и в клетках запускается программа апоптоза, «альтруистического самоубийства» клеток. Хотя граница Хейфлика и считается основной причиной уменьшения уровня клеточного деления, даже стволовые клетки, в которых эта граница отсутствует, становятся менее активными, замедляют своё деление и не так часто дифференцируются в соматические клетки. Серьёзной причиной, тормозящей прогресс в области изучения старения клеток, является то, что исследования в этой сфере в основном выполняются на большой совокупности клеток в один из моментов роста клеточной популяции. Однако такой подход не позволяет проследить всю последовательность изменений, начиная с их возникновения в отдельных клетках и заканчивая экспансией этих изменений в клеточной популяции. См. также:
Генеалогия клеток
Человек и мышь — виды млекопитающих, резко контрастные по продолжительности жизни и частоте возникновения онкологических заболеваний, которая у грызунов на несколько порядков выше, чем у человека. Сравнение изменений фенотипа клеток у человека и мыши позволяет получать новые данные о генетических процессах, ведущих к старению и трансформации. Анализ видеозаписей компьютерной видеосъемки живых клеточных культур, произведённых для изучения изменений фенотипа отдельных клеток и их потомств, позволил составлять клеточные генеалогии, напоминающие родословные в генетике человека. Клетки-родоначальницы и их потомки в течение ряда поколений изучались по многим признакам. Учитывалось до 20 параметров, таких как площадь и форма клетки, митозы и клеточная гибель, аномальные митозы, размер и количество ядер, клазматоз, скорость и характер перемещений клеток и др.
Кожа и соединительная ткань
Формирование перекрёстных связей между молекулами коллагена, основного структурного межклеточного белка организма млекопитающих, и кальцификация гладких мышц и стенок сосудов, увеличивает жёсткость соединительной ткани. При этом одновременно проходит декальцификация костей скелета, в результате чего кости становятся тоньше, менее надёжными и менее крепкими. Из-за истончения хрящей позвоночника уменьшается длина тела.
Нервная система
В большинстве тканей происходит атрофия клеток и даже целых структур, особенно это заметно в некоторых тканях, которые медленно восстанавливаются, в первую очередь центральной нервной системы. Хотя даже количество периферийных нервных волокон человека уменьшается на 20 процентов к 90 годам, больше всего страдают клетки коры головного мозга. Эта потеря нейронов — главная причина снижения умственных способностей пожилых людей, хотя сказывается и некоторый эффект снижения поступления кислорода. Увеличивается также накопление прионподобных белковых скоплений, приводящих у человека к деменции, а также болезням Альцгеймера и Паркинсона.
Накопление веществ, не выполняющих положительной роли
Также для многих тканей характерны отложения инертных и потенциально опасных веществ. Например, пигмент липофусцин, отсутствующий в молодости, в старости составляет до 3 % массы сердечной мышцы. Известны отложения и в кровеносных сосудах — атеросклероз.
Также заметны изменения в эндокринной системе, которая замедляет ответ на изменения внешней среды, в результате организм становится более чувствительным к каким-либо неблагоприятным факторам (стресс).
Иммунная система
Из-за старения иммунной системы увеличивается вероятность аутоиммунных реакций при общем снижении активности тимус-зависимой подсистемы. В результате как увеличивается вероятность развития опухолей из раковых клеток, так и возникает риск аутоиммунных заболеваний.
Генетика старения
Генетика старения эукариот
Во время старения клетки проходят через ряд генетических изменений, которые тем или иным образом влияют на ход процесса старения. Исследование старения эукариот были проведены с помощью целого ряда модельных организмов, таких как домовая мышь (Mus musculus), плодовая муха (Drosophila melanogaster), нематоды Caenorhabditis elegans, гриб Podospora anserina и дрожжи Saccharomyces cerevisiae. Исследования этих организмов выявили присутствие как минимум двух путей, которые активируются во время старения.
| Podospora | Saccharomyces | Caenorhabditis | Drosophila | Мышь |
|---|---|---|---|---|
| grisea | LAG1 | daf-2 | sod1 | Prop-1 |
| LAC1 | age-1/daf-23 | cat1 | p66shc | |
| RAS1 | daf-18 | mth | mclk1 | |
| RAS2 | akt-1/akt-2 | |||
| PHB1 | daf-16 | |||
| PHB2 | daf-12 | |||
| CDC7 | ctl-1 | |||
| BUD1 | old-1 | |||
| RTG2 | spe-26 | |||
| RPD3 | clk-1 | |||
| HDA1 | mev-1 | |||
| SIR2 | ||||
| SIR4-42 | ||||
| UTH4 | ||||
| YGL023 | ||||
| SGS1 | ||||
| RAD52 | ||||
| FOB1 |
В одном из этих путей принимает участие продукт гена SIR2, НАД±зависимая деацетилаза гистонов. В дрожжах белок Sir2 необходим для репрессии генов в трёх локусах (местоположениях): локусе спаривания дрожжей, генах теломер и рибосомальной рДНК. В некоторых видах дрожжей гомологическая рекомбинация между повторами рДНК приводит к исключению повторов рДНК и созданию экстрахромосомной кольцевой рРНК (ЭКР или ERC, от англ. Exctachtomosomal rRNA circles). Эти ЭЦР реплицируются и преимущественно аккумулируются в материнской клетке во время деления, и вызывают клеточное старение за счёт конкурентного связывания с важными факторами. ЭЦР не наблюдались в других видах дрожжей (которые также проявляют репликативное старение) и в высших организмах, таких как человек. Экстрахромосомная кольцевая ДНК (экДНК, eccDNA) была найдена у червей, мух и человека. Роль экДНК в старении, если она и есть, неизвестна.
Несмотря на отсутствие связи между кольцевой ДНК и старением высших организмов, дополнительные копии гомологов гена Sir2 способны к увеличению продолжительности жизни как червей, так и мух. Механизмы, которыми гомологи Sir2 у высших организмов регулируют продолжительность жизни, остаются неясными, но было установлено, что белок человека SIRT1 деацетилирует p53, Ku70 и семейство факторов транскрипции forkhead. SIRT1 также может регулировать другие ацетилированные белки, такие как CBP/p300, и может деацетилировать некоторые аминокислоты гистонов.
RAS1 и RAS2 также влияют на старение в дрожжах и имеют человеческие гомологи. Было показано, что повышенная экспрессия RAS2 увеличивает продолжительность жизни дрожжей.
Несколько других генов регулирует старение в дрожжах, увеличивая сопротивление окислительному стрессу. Супероксиддисмутаза, белок, который защищает клетку против эффектов активных форм кислорода (АФК) митохондрий, может замедлить старение дрожжей, если они сильно экспрессируются на протяжении стационарной фазы.
В процессах регуляции старения высших организмов важную роль играет инсулин-подобный ростовой фактор IGF-1. Мутации, которые влияют на инсулин-подобный сигнальный путь в червях, мухах и мышах, часто связаны с увеличенной продолжительностью жизни. Этот путь угнетается при ограничении потребления калорий, и в свою очередь влияет на продолжительность жизни через механизм, зависимый от белков p53/p21/Akt.
В дрожжах активность Sir2 регулируется никотинамидазой PNC1. Производство PNC1 увеличивается на уровне транскрипции в условиях стресса, например, во время низкокалорийной диеты, теплового или осмотического шока. Перерабатывая никотинамид в ниацин, никотинамидаза использует никотинамид, который ингибирует активность Sir2. Никотинамидаза найдена и у человека, где она известна как PBEF и, возможно, выполняет подобную функцию, а секретированная форма PBEF, известная как висфатин, возможно, помогает регулировать уровень инсулина в сыворотке. Неизвестно, однако, эти ли механизмы также существуют у человека из-за значительных различий в физиологии мыши и человека.
Было показано, что активность Sir2 возрастает при условиях ограничения калорий в диете мышей. В связи с отсутствием доступной глюкозы в клетках создаётся больше свободного NAD+, что приводит к активации Sir2. Резвератрол, полифенол найденный в некоторых фруктах, увеличивает продолжительность жизни дрожжей, червей и мух за счёт активизации деятельности Sir2 и имитации эффекта низкокалорийной диеты.
По некоторым данным, процессу старения могут благоприятствовать и флуктуации в экспрессии многих генов. Индивидуальные, генетически идентичные клетки могут иметь существенно различные ответы на внешние стимулы и заметно разную продолжительность жизни, указывая, что эпигенетические факторы играют важную роль в экспрессии генов и старении.
Генетика старения бактерий
Намного меньше известно о старении бактерий, несмотря на их более простую структуру и удобство наблюдения. Среди бактерий лучше известны изменения, которые происходят во время условного старения (хронологического старения в стационарной фазе) бактерии E. coli.
Большая часть генетически контролируемых изменений во время условного старения E. coli происходит из-за изменения в уровне экспрессии сигма-фактора σs, который отвечает за экспрессию генов, связанных с ремонтом повреждённых белков, аналогично гену C. elegans daf-16 и геном дрожжей RAS/PKA. σs конкурирует с другим сигма-фактором, σ70, который отвечает за рост бактерии, и никогда не экспрессируется в «полную силу». Таким образом, бактерия продолжает ограниченный рост даже в условиях стационарной фазы, которая даёт ей возможность быстро возобновить рост, если условия изменяются, но недостатком является невозможность σs справиться со значительным оксидативным стрессом. Таким образом, остаточная активность σ70 в стационарной фазе является примером так называемой антагонистической плейотропии (см. ниже), типа генетической системы, которая развивается за счёт позитивного эффекта на одних стадиях жизни, несмотря на негативный эффект на других, более редких, стадиях.
Причины старения
История исследования
Первые попытки научного объяснения старения начались в конце XIX века. В одной из первых работ Вейсман предложил теорию происхождения старения как свойства, которое возникло в результате эволюции. Согласно Вейсману, «не стареющие организмы не только не являются полезными, они вредны, потому что занимают место молодых», что, согласно Вейсману, должно было привести эволюцию к возникновению старения.
Важным шагом в исследовании старения был доклад профессора Питера Медавара перед Лондонским королевским обществом в 1951 году под названием «Нерешённая проблема биологии». В этой лекции он подчеркнул, что животные в природе редко доживают до возраста, когда старение становится заметным, таким образом эволюция не могла оказывать влияние на процесс развития старения. Эта работа положила начало целой серии новых исследований.
На протяжении следующих 25 лет исследования имели преимущественно описательный характер. Тем не менее, начиная с конца 70-х годов, возникает большое количество теорий, которые пытались объяснить старение. Например, в известном обзоре литературы по этому вопросу, опубликованном Калебом Финчем в 1990 году, насчитывалось около 4 тыс. ссылок. Только в конце 1990-х годов ситуация начала проясняться, и большинство авторов начали приходить к общим выводам.
Все теории старения можно условно разделить на две большие группы: эволюционные теории и теории, основанные на случайных повреждениях клеток. Первые считают, что старение является не необходимым свойством живых организмов, а запрограммированным процессом. Согласно им, старение развилось в результате эволюции из-за некоторых преимуществ, которые оно даёт целой популяции. В отличие от них, теории повреждения предполагают, что старение является результатом природного процесса накопления повреждений со временем, с которыми организм старается бороться, а различия старения у разных организмов является результатом разной эффективности этой борьбы. Сейчас последний подход считается установленным в биологии старения. Тем не менее, некоторые исследователи всё ещё защищают эволюционный подход, а некоторые другие совсем игнорируют деление на эволюционные теории и теории повреждений. Последнее утверждение является частично результатом смены терминологии: в некоторых работах последнего времени термин «эволюционные теории» ссылается не на теории «запрограммированного старения», которые предлагают эволюционное возникновение старения как полезного явления, а на подход, который описывает почему организмы должны стареть в противоположность вопросу о биохимических и физиологических основах старения.
Почему возникает старение
Эволюционно-генетический подход
Гипотеза, которая легла в основу генетического подхода, была предложена Питером Медаваром в 1952 году и известна сейчас как «теория накопления мутаций» (англ. Mutations accumulation theory). Медавар заметил, что животные в природе очень редко доживают до возраста, когда старение становится заметным. Согласно его идее, аллели, которые проявляются на протяжении поздних периодов жизни и которые возникают в результате мутаций зародышевых клеток, подвергаются довольно слабому эволюционному давлению, даже если в результате их действия страдают такие свойства, как выживание и размножение. Таким образом, эти мутации могут накапливаться в геноме на протяжении многих поколений. Тем не менее, любая особь, которая сумела избежать смерти на протяжении долгого времени, испытывает на себе их действие, что проявляется как старение. То же самое верно и для животных в защищённых условиях.
В дальнейшем, в 1957 году Д. Вильямс предположил существование плейотропных генов, которые имеют разный эффект для выживания организмов на протяжении разных периодов жизни, то есть они полезны в молодом возрасте, когда эффект естественного отбора сильный, но вредны позднее, когда эффект естественного отбора слабый. Эта идея сейчас известна как «антагонистическая плейотропия» (англ. Antagonistic pleiotropy).
Вместе эти две теории составляют основу современных представлений о генетике старения. Тем не менее, идентификация ответственных генов имела лишь ограниченный успех. Свидетельства о накоплении мутаций остаются спорными, тогда как свидетельства наличия плейотропных генов сильнее, но и они недостаточно обоснованы. Примерами плейотропных генов можно назвать ген теломеразы у эукариот и сигма-фактор σ70 у бактерий. Хотя известно много генов, которые влияют на продолжительность жизни разных организмов, других чётких примеров плейотропных генов всё ещё не обнаружено.
Эволюционно-физиологический подход
Теория антагонистической плейотропии предсказывает, что должны существовать гены с плейотропным эффектом, естественный отбор которых и приводит к возникновению старения. Несколько генов с плейотропным эффектом на разных стадиях жизни действительно найдены — сигма-70 у E. coli, теломераза у эукариот, но непосредственной связи со старением показано не было, тем более не было показано, что это типичное явление для всех организмов, ответственное за все эффекты старения. То есть эти гены могут рассматриваться лишь как кандидаты на роль генов, предсказанных теорией. С другой стороны, ряд физиологических эффектов показаны без определения генов, ответственных за них. Часто мы можем говорить о компромиссах, аналогичных предсказанным теорией антагонистической плейотропии, без чёткого определения генов, от которых они зависят. Физиологическая основа таких компромиссов заложена в так называемой «теории одноразовой сомы» (англ. Disposable soma theory). Эта теория задаётся вопросом, как организм должен распорядиться своими ресурсами (в первом варианте теории речь шла только о энергии) между поддержкой, ремонтом сомы и другими функциями, необходимыми для выживания. Необходимость компромисса возникает из-за ограниченности ресурсов или необходимости выбора лучшего пути их использования.
Поддержание тела должно осуществляться только настолько, насколько это необходимо на протяжении обычного времени выживания в природе. Например, поскольку 90 % диких мышей умирает на протяжении первого года жизни (преимущественно от холода), инвестиции ресурсов в выживание на протяжении дольшего времени будут касаться только 10 % популяции. Трёхлетняя продолжительность жизни мышей полностью достаточна для всех потребностей в природе, но с точки зрения эволюции ресурсы следует тратить, например, на улучшение сохранения тепла или размножения вместо борьбы со старостью. Таким образом, продолжительность жизни мыши наилучшим образом отвечает экологическим условиям её жизни.
Теория «одноразового тела» делает несколько допущений, которые касаются физиологии процесса старения. Согласно этой теории, старение возникает в результате неидеальных функций ремонта и поддержки соматических клеток, которые адаптированы для удовлетворения экологических потребностей. Повреждения, в свою очередь, являются результатом стохастических процессов, связанных с жизнедеятельностью клеток. Долголетие контролируется за счёт контроля генов, которые отвечают за эти функции, а бессмертие генеративных клеток, в отличие от соматических, является результатом больших затрат ресурсов и, возможно, отсутствия некоторых источников повреждений.
Как возникает старение
Молекулярные механизмы
Существуют свидетельства нескольких важнейших механизмов повреждения макромолекул, которые обычно действуют параллельно один другому или зависят один от другого. Вероятно, любой из этих механизмов может играть доминирующую роль при определённых обстоятельствах.
Во многих из этих процессов важную роль играют активные формы кислорода (в частности свободные радикалы), набор свидетельств о их влиянии был получен достаточно давно и сейчас известен под названием «свободнорадикальная теория старения». Сегодня, тем не менее, механизмы старения намного более детализированы. Так, например, обнаружено что характер метилирования ДНК в геноме тесно связан с хронологическим возрастом
Теория соматических мутаций
Многие работы показали увеличение с возрастом числа соматических мутаций и других форм повреждения ДНК, предлагая репарацию (ремонт) ДНК в качестве важного фактора поддержки долголетия клеток. Повреждения ДНК типичны для клеток, и вызываются такими факторами как жёсткая радиация и активные формы кислорода, и потому целостность ДНК может поддерживаться только за счёт механизмов репарации. Действительно, существует зависимость между долголетием и репарацией ДНК, как это было продемонстрировано на примере фермента поли-АДФ-рибоза-полимеразы-1 (PARP-1), важного игрока в клеточном ответе на вызванное стрессом повреждение ДНК. Более высокие уровни PARP-1 ассоциируются с большей продолжительностью жизни.
Накопление изменённых белков
Также важен для выживания клеток кругооборот белков, для которого критично появление повреждённых и лишних белков. Окисленные белки являются типичным результатом влияния активных форм кислорода, которые образуются в результате многих метаболических процессов клетки и часто мешают корректной работе белка. Тем не менее, механизмы репарации не всегда могут распознать повреждённые белки и становятся менее эффективными с возрастом за счёт снижения активности протеасомы. В некоторых случаях белки являются частью статических структур, таких как клеточная стенка, которые не могут быть легко разрушены. Кругооборот белков зависит также и от белков-шаперонов, которые помогают белкам получать необходимую конформацию. С возрастом наблюдается снижение репарирующей активности, хотя это снижение может быть результатом перегрузки шаперонов (и протоасомы) повреждёнными белками.
Существуют свидетельства, что накопление повреждённых белков действительно происходит с возрастом и может отвечать за такие ассоциированные с возрастом болезни как болезнь Альцгеймера, болезнь Паркинсона и катаракта.
Митохондриальная теория
Митохондриальная теория старения впервые была предложена в 1978 году (митохондриальная теория развития, старения и злокачественного роста). Суть её заключается в том, что замедление размножения митохондрий в высокодифференцированных клетках вследствие дефицита кодируемых в ядре митохондриальных белков создает условия для возникновения и селективного отбора дефектных делеционных мтДНК, увеличение доли которых постепенно снижает энергетическое обеспечение клеток. В 1980 году была предложена радикальная митохондриальная теория старения. В настоящее время накопилось много данных[где?] свидетельствующих о том, что свободные радикалы не являются причиной естественного старения. Эти данные не опровергают митохондриальную теорию старения (1978 г.), которая не опирается на свободные радикалы, но доказывают ложность радикального варианта митохондриальной теории старения (1980 г.).
Важность связи между молекулярным стрессом и старением была предположена, основываясь на наблюдениях за эффектом накопления мутаций в митохондриальной ДНК (мтДНК). Эти данные были подкреплены наблюдением увеличения с возрастом числа клеток, которым не хватает цитохром-с-оксидазы (COX), что ассоциировано с мутациями мтДНК. Такие клетки часто имеют нарушения в производстве АТФ и клеточном энергетическом балансе.
Утрата теломер
Во многих клетках человека утрата способности клеток к делению связана с утратой теломер на концах хромосом, которые утрачиваются после определённого количества делений. Это происходит из-за отсутствия фермента теломеразы, который обычно экспрессуется только у зародышевых и стволовых клеток. Теломераза позволяет им непрерывно делиться, формируя ткани и органы. У взрослых организмов теломераза экспрессируется в клетках, которые должны часто делиться, однако большинство соматических клеток её не производят. Теломеразу считают ключом к клеточному бессмертию, «источником юности». Этот фермент позволяет клеткам быстро размножаться без старения.
Поскольку на настоящий момент неизвестно, насколько разрушение теломер влияет на процесс старения, основные исследования направлены на процессы сохранения целостности ДНК и в особенности её теломерных участков. Майкл Фоссел в одном из интервью предположил, что лечение теломеразой может использоваться не только для борьбы с раком, но даже для борьбы со старением человеческого организма и таким образом для увеличения продолжительности жизни. Он считает, что уже в ближайшем десятилетии будут проведены первые испытания теломеразных методов увеличения продолжительности жизни человека.
Недавно было обнаружено, что окислительный стресс (чрезмерное выделение активных форм кислорода) также может иметь влияние на утрату теломер, значительно ускоряя этот процесс в определённых тканях.
Эпигенетическая теория старения
Клетки со временем медленно теряют маркеры репрессированного хроматина, что может быть связано с дифференцировкой клеток в организме. Утрата маркеров репрессии рано или поздно должна приводить к дерепрессии дремлющих транспозонов, соответственно, к росту количества вызванных ими повреждений ДНК с последующей активацией клеточных систем репарации ДНК. Последние, помимо участия в восстановлении ДНК, вызывают и несанкционированные рекомбинации в теломерах. Также не исключено, что рекомбиназы транспозонов могут непосредственно инициировать подобные рекомбинации. В результате протяженные участки теломерной ДНК преобразуются в кольца и теряются, а теломеры укорачиваются на длину утраченной кольцевой ДНК. Данный процесс ускоряет утрату теломерной ДНК в десятки раз, а последующий апоптоз большинства клеток и предопределяет старение как биологическое явление. Предложенная теория является альтернативой гипотезе о генетически запрограммированном старении и гипотезе о старении как следствии накопления ошибок и повреждений, объясняет механизм ускорения утраты теломер в случае окислительного стресса и повреждений ДНК, а также взаимосвязь старения и возникновения опухолей.
Системные и сетевые механизмы
На первых этапах исследования старения, многочисленные теории рассматривались как конкурирующие в пояснении эффекта старения. Тем не менее, сегодня считается, что многие механизмы повреждения клеток действуют параллельно, и клетки также должны тратить ресурсы на борьбу со многими механизмами. Для исследования взаимодействия между всеми механизмами борьбы с повреждениями был предложен системный подход к старению, который пытается одновременно принять во внимание большое количество таких механизмов. Более того, этот подход может чётко разделить механизмы, которые действуют на разных стадиях жизни организма. Например, постепенное накопление мутаций в митохондриальной ДНК часто приводит к накоплению активных форм кислорода и снижению производства энергии, что в свою очередь приводит к увеличению скорости повреждения ДНК и белков клеток.
Другой аспект, который делает системный подход привлекательным, это понимание разницы между разными типами клеток и тканей организма. Например, клетки, которые активно делятся, с большей вероятностью пострадают от накопления мутаций и утраты теломер, чем дифференцированные клетки. В то же время необходимо уточнить, что данный тезис не относится к быстро и многократно делящимся трансформированным и опухолевым клеткам, которые не утрачивают теломеры и не накапливают мутации. Дифференцированные клетки с большей вероятностью пострадают от повреждения белков, чем клетки, которые быстро делятся и «разбавляют» повреждённые белки вновь синтезированными. Даже если клетка теряет способность к пролиферации за счёт процессов старения, баланс механизмов повреждения в ней сдвигается.
Популяционный подход
Другим подходом к изучению старения являются исследования популяционной динамики старения. Все математические модели старения можно примерно разбить на два главных типа: модели данных и системные модели. Модели данных — это модели, которые не используют и не пытаются пояснить какие-либо гипотезы о физических процессах в системах, для которых эти данные получены. К моделям данных относятся, в частности, и все модели математической статистики. В отличие от них, системные модели строятся преимущественно на базе физических законов и гипотез о структуре системы, главным в них является проверка предложенного механизма.
Первым законом старения является закон Гомпертца, который предлагает простую количественную модель старения. Этот закон даёт возможность разделить два типа параметров процесса старения. Исследования отклонения закона старения от кривой Гомпертца могут дать дополнительную информацию относительно конкретных механизмов старения данного организма. Самый известный эффект такого отклонения — выход смертности на плато в позднем возрасте вместо экспоненциального роста, наблюдавшийся во многих организмах. Для пояснения этого эффекта было предложено несколько моделей, среди которых вариации модели Стрелера-Милдвана и теории надёжности.
Системные модели рассматривают много отдельных факторов, событий и явлений, которые непосредственно оказывают влияние на выживание организмов и рождение потомства. Эти модели рассматривают старение как баланс и перераспределение ресурсов как в физиологическом (в течение жизни одного организма), так и в эволюционном аспектах. Как правило, особенно в последнем случае, речь идёт о распределении ресурсов между непосредственными затратами на рождение потомства и затратами на выживание родителей.
Клеточный ответ на старение
Важным вопросом старения на уровне клеток и ткани является клеточный ответ на повреждения. Из-за стохастической природы повреждений отдельные клетки стареют, например в связи с достижением границы Хейфлика, быстрее остальных клеток. Такие клетки потенциально могут угрожать здоровью всей ткани. В наибольшей мере такая угроза проявляется среди стволовых клеток, у которых происходит быстрое деление, таких как клетки костного мозга или эпителия кишечника, в связи с большим потенциалом таких тканей в создании мутантных, возможно раковых, клеток. Известно, что именно клетки этих тканей быстро отвечают на повреждения инициацией программы апоптоза. Например, даже низкие дозы радиации (0,1 Gy) вызывают апоптоз в клетках эпителия кишечника, а даже слабый химический стресс вызывает апоптоз стволовых клеток старых мышей.
Как правило, в таких тканях массовый апоптоз является признаком возрастания числа повреждений клеток. С другой стороны, в других тканях ответом на возрастание уровня повреждений может быть арест клеток на определённой стадии клеточного цикла для прекращения деления. Баланс между апоптозом и арестом повреждённых клеток наиболее важен как компромисс между старением и раком. То есть, или организм должен убить повреждённые клетки, или дать им возможность существовать, увеличивая риск возникновения рака. Таким образом, p53 и сокращение теломер, важные факторы в вызывании апоптоза клеток, могут рассматриваться как пример антагонистической плейотропии, как было указано выше.
Подводя итог, по современным представлениям, клетка стареет в результате накопления повреждений. Скорость этого накопления определяется, в первую очередь, генетически определёнными затратами на ремонт и поддержку клеточных структур, которые в свою очередь определяются организмом для удовлетворения своих экологических потребностей. Долгоживущие организмы имеют большие затраты (иногда более длительный метаболизм), что приводит к более медленному накоплению повреждений. Для борьбы с риском, который представляют собой повреждённые клетки, организм создал систему механизмов для борьбы с ними, которые часто включают второй ряд компромиссов.
Политическая борьба против старения
В июле 2012 сначала в России, а затем в США, Израиле и Нидерландах было объявлено о начале создания политических партий продления жизни. Эти партии нацелены на оказание политической поддержки научно-технической революции, идущей сейчас в сфере продления жизни и обеспечении максимально быстрого и одновременно безболезненного перехода общества на следующий этап своего развития-с радикальным увеличением продолжительности человеческой жизни, омоложением и остановкой старения для того, чтобы большинство живущих в настоящее время людей успели воспользоваться достижениями науки и увеличить свою жизнь. Ведущие мировые ученые в области старения считают, что старение должно официально быть включено в список болезней и болезненных синдромов Всемирной организации здравоохранения, о чём в адрес ВОЗ было направлено коллективное письмо по результатам 3-й Международной конференции «Генетика старения и долголетия», которая проходила в Сочи в апреле 2014 года.
См. также
- Геропротекторы
- Иммортализм
- Клото
- Максимальная продолжительность жизни
- Изменения метилирования ДНК при старении
- Популяционная динамика старения
- Предел Хейфлика
- Пренебрежимое старение
- Прогерия
- Закон смертности Гомпертца — Мейкхама
- Теории эволюции старения
- Теломераза (Старение)
- Смерть
- Старение бактерий
- Старение человека
- Феноптоз
- Физиология старения млекопитающих
- Хронология исследования старения
Литература
Энциклопедии
- Microsoft Encarta: Aging, Life Span (Англ. язык, требует предоплату)
- Encyclopaedia Britannica: Aging, Life Span (Англ. язык, требует предоплату)
Общие научные обзоры
- Mechanisms of Aging (Англ. язык, описание механизмов/теорий старения.)
- Aging Series edited by Christopher B. Newgard and Norman E. Sharpless (2013) (Англ. язык, описание молекулярных механизмов старения.)
- Jonathan Silvertown (2013) The Science of Life Span and Aging. University of Chicago Press, Chicago, ISBN 978-0-226-75789-6 (Англ. язык, популярное описание механизмов/теорий старения.)
- (2015). Molecular and Cellular Biology of Aging (Англ. язык, описание механизмов старения.)
- Victoria, V., Gaur, M. G., & Amaro-Ortiz, A. (2017). Mesenchymal Stem Cells Secretory Responses: Senescence-Messaging System and Immunomodulation Perspective. Frontiers in genetics, 8, 220. doi:10.3389/fgene.2017.00220 PMC 5742268 В статье подробно описаны типы старения клеток и молекулярные механизмы различных типов старения
- Анисимов В.Н. (2008). Молекулярные и физиологические механизмы старения скачать книгу PDF
Отдельные теории
- Theodore Goldsmith. The Evolution of Aging (англ.). (Англ. язык, статья, защищающая «Теорию запрограммированной смерти» или «Эволюционную теорию» возникновения старения.)
- Evolutionary Theories of Aging and Longevity (Англ. язык, обзор эволюционного подхода к старению.)
- Gavrilov, L. A. Reliability Theory of Aging and Longevity : [англ.] : [арх. 1 июля 2004] / L. A. Gavrilov, N. S. Gavrilova // Journal of theoretical Biology. — 2001. — P. 527–545. — doi:10.1006/jtbi.2001.2430.
- Leonard Hayflick. Biological Aging Is No Longer an Unsolved Problem // Annals of the New York academy of Sciences. — 2007. — Т. 1100. — С. 1—13. Архивировано 6 июля 2007 года. (Англ. язык, статья против эволюционной теории, написанная Леонардом Хейфликом, открывателем предела Хейфлика, бывшим главой Геронтологического Института США и основателем Национального Института Старения США.)
- Thomas Nystrom. Conditional senescense in bacteria: death of the immortals (англ.) // Molecular Microbiology : journal. — 2003. — Vol. 48. — doi:10.1046/j.1365-2958.2003.03385.x.
- Peter Launa et al. Yeast as a model for chronological and reproductive aging – A comparison (англ.) // Experimental Gerontology : journal. — 2006. — Vol. 41, no. 12. — P. 1208—1212. (недоступная ссылка) (Англ. язык, обзор старения дрожжей.)
- Московский ученый обнаружил причину старения человека
- Эпигенетическая теория старения (недоступная ссылка с 13-05-2013 [3662 дня])
Организации, списки ресурсов
- eternalmind.ru (Рус. язык, Новости Современной Геронтологии. (наука о старении)
- Biologicalgerontology.com (Англ. язык, ресурсы о биологии старения и увеличения продолжительности жизни.)
- Unraveling the Secrets of Human Longevity (Англ. язык, ресурсы о биологии старения и увеличения продолжительности жизни.)
- Senescence.info (Англ. язык, ресурсы о биологии старения.)
- Cellular Senescence Information Center (недоступная ссылка с 13-05-2013 [3662 дня] — история) (Англ. язык, ресурсы о клеточном старении.)
- American Academy of Anti-Aging Medicine (Англ. язык, Американская Ассоциация Медицины против Старения.)
- American Federation for Aging Research (Англ. язык, Американская Ассоциация Исследования Старения.)
- Aging Research Centre (ARC) (Англ. язык, Центр Исследования Старения.)
Базы данных
- AnAge (Англ. язык, База данных по старению и продолжительности жизни животных), распространяется по лицензии Creative Commons
Динамические изображения
| В библиографических каталогах |
|---|
| Вопросы |  |
|
|---|---|---|
| Журналы | ||
| Ресурсы | ||
| Люди | ||
| Организации | ||
Категория «Продление жизни» | ||