
| Пресмыкающиеся | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
  | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен:
Царство:
Подцарство:
Без ранга:
Без ранга:
Тип:
Подтип:
Инфратип:
Надкласс:
Клада:
Клада:
Класс:
Пресмыкающиеся
|
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Reptilia Laurenti, 1768 | |||||||||||||||||||||||||||||||||||||||||
| Современные отряды | |||||||||||||||||||||||||||||||||||||||||
|
Геохронология появился 312 млн лет
◄
Наше время
◄
Мел-палеогеновое вымирание ◄
Триасовое вымирание
◄
Массовое пермское вымирание
◄
Девонское вымирание
◄
Ордовикско-силурийское вымирание
◄
Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
Пресмыка́ющиеся, или репти́лии (лат. Reptilia), — традиционно выделяемый класс преимущественно наземных позвоночных животных из клады амниот, включающий современных черепах, крокодилов, клювоголовых и чешуйчатых. Пресмыкающиеся являются парафилетической группой, так как дали начало птицам, но не включают их самих. В качестве монофилетической альтернативы класса выделяется клада завропсид, или рептилий в пересмотренном составе, в которую включают представителей традиционного класса пресмыкающихся (кроме зверообразных) и птиц.
В XVIII—XIX веках вместе с амфибиями объединялись в группу гадов — холоднокровных наземных позвоночных. Традиционно к этой группе относили различных позвоночных, согласно изначальным представлениям похожих по своей организации на современных рептилий (например, немаммальных синапсид — предков и родственников современных млекопитающих). Однако в настоящее время ряд вопросов о физиологии многих вымерших групп организмов остаётся открытым, а данные об их генетических и эволюционных связях не поддерживают подобного рода классификацию.
Рептилии пережили расцвет в эпоху мезозоя, когда они доминировали на суше, в море и в воздухе. Крупнейшие наземные животные принадлежали к динозаврам. В конце мелового периода большая часть рептилий, особенно крупного размерного класса, вымерла. Однако бытующее мнение, что современные пресмыкающиеся являются лишь разрозненными остатками того древнего мира, неверно. Выделяется около 11 940 рецентных видов пресмыкающихся, большая часть которых относится к отряду чешуйчатых, на территории России обитает 77 видов[обновить данные]. Небольшие хищные динозавры дали начало процветающей в настоящее время группе животных — птицам, причём многие приспособления, обусловившие эволюционный успех этой группы, появились ещё у её архозавровых предков, представлявших собой специализированную группу диапсид (теплокровность, теплоизолирующий покров тела — перья, развитый головной мозг, и так далее).
Эволюция пресмыкающихся
Предки пресмыкающихся — рептилиоморфы — известны со среднего карбона. Ранее первыми рептилиями считали представителей группы котилозавров, в которую включали различные малосвязанные таксоны рептилиоморф. Продвинутые рептилиоморфы — амниоты — сразу разделились на две ветви: синапсиды и завропсиды. К синапсидам относятся млекопитающие и их вымершие родственники, ранее известные как «зверообразные рептилии» (с точки зрения кладистической таксономии пресмыкающимися не являются). К завропсидам, в свою очередь, относятся пресмыкающиеся и филогенетически вложенные в их состав птицы. В конце пермского периода оформляются основные эволюционные линии рептилий, включая архозавроморф и лепидозавроморф. В триасе появляются первые динозавры, крокодиломорфы, завроптеригии, первые черепахи и прочие группы. В мезозойской эре наступает расцвет пресмыкающихся, среди представителей наблюдается наибольшее многообразие. Происходит освоение морских и речных водоёмов, а также воздушного пространства. В юрском периоде одна из ветвей тероподовых динозавров — манирапторы — эволюционирует в первых птиц (авиал). Последняя группа — змеи — сформировалась в меловом периоде.
В конце мелового периода происходит резкое сокращение количества видов пресмыкающихся. Однозначно указать причины вымирания современная наука пока не может.
Происхождение
Произошли рептилии примерно 310—320 миллионов лет назад, в болотах позднего каменноугольного периода, когда из продвинутых рептиломорфов стали выделяться другие виды.
Самым старым известным животным, которое могло быть амниотой, является Casineria (хотя, возможно, это был темноспондил). Обнаружена целая серия следов из ископаемых пластов Новой Шотландии, датированных 315 миллионов лет назад; видны типичные для рептилий пальцы ног и отпечатки чешуи. Эти следы приписываются Hylonomus, старейшей из известных рептилий. Это было маленькое, похожее на ящерицу животное, от 20 до 30 сантиметров (от 7,9 до 11,8 дюйма) в длину, с многочисленными острыми зубами, указывающими на питание в основном насекомыми. Другие примеры включают Westlothiana (на данный момент считается лишь рептилиоморфом, а не настоящей амниотой) и Paleothyris, оба похожего телосложения.
Однако иногда микрозавров считали настоящими рептилиями, поэтому возможно более раннее происхождение пресмыкающихся.
Дальнейшая история
Самые ранние амниоты, в том числе первые рептилии (те амниоты, которые ближе к современным рептилиям, чем к млекопитающим), были в значительной степени затенены более крупными четвероногими, такими как Cochleosaurus, и оставались небольшой, незаметной частью фауны до краха каменноугольных тропических лесов. Этот внезапный коллапс затронул несколько больших групп. Примитивные четвероногие были особенно затронуты, многие исчезли, в то время как рептилии жили лучше, будучи экологически адаптированными к более засушливым условиям. Примитивным четвероногим, как и современным амфибиям, было необходимо вернуться в воду, чтобы отложить яйца; напротив, амниоты, как и современные рептилии, яйца которых имеют скорлупу, позволяющую откладывать их на сушу, лучше приспособились к новым условиям жизни. Амниоты занимали новые ниши быстрее, чем до коллапса, и гораздо быстрее, чем примитивные четвероногие. Они приобрели новые стратегии питания, в том числе травоядные и плотоядные (ранее они были только насекомоядными и рыбоядными). С этого момента рептилии доминировали в пищевых цепях и отличались большим разнообразием, чем примитивные четвероногие, что подготовило почву для мезозоя (известного как эпоха рептилий). Одной из самых известных ранних рептилий является Mesosaurus, род из ранней перми, который обитал в воде, питаясь рыбой.
Исследование разнообразия рептилий в каменноугольном и пермском периодах, проведённое в 2021 году, предполагает гораздо более высокую степень разнообразия, чем считалось ранее, сопоставимую или даже превышающую таковую у синапсид. Таким образом началась «Первая эпоха рептилий».
Анапсиды, синапсиды, диапсиды и завропсиды
Традиционно предполагалось, что первые рептилии сохранили унаследованный от предков анапсидный череп, имеющий отверстия только для глаз и ноздрей. Открытие синапсидоподобных отверстий (см. ниже) в черепе нескольких представителей Parareptilia (клады, содержащей большинство амниот, традиционно называемых «анапсидами»), включая лантанозухоидов, миллереттид, болозавридов, некоторые никтеролетериды, некоторые проколофоноиды и, по крайней мере, некоторые мезозавры сделало его более двусмысленным, и в настоящее время неясно, имел ли предковый амниот анапсидоподобный или синапсидоподобный череп. Этих животных традиционно называют «анапсиды», и они образуют парафилетическую группу, из которого произошли другие группы. Очень скоро после того, как появились первые амниоты, отделились животные, названные Synapsida. Это «амниоты, похожие на млекопитающих», или предки млекопитающих, которые позже дали начало настоящим млекопитающим. Вскоре после этого другая группа развила аналогичную черту, на этот раз с двойным отверстием за каждым глазом, за что они получили название Diapsida («две арки»). Функция отверстий в этих группах заключалась в том, чтобы облегчить череп и дать место для движения мышц челюсти, что позволило сделать более сильный укус.
Традиционно считалось, что черепахи являются выжившими парарептилиями на основании их анапсидной структуры черепа, которая считалась примитивной чертой. Обоснование этой классификации оспаривалось, некоторые утверждали, что черепахи являются диапсидами, которые развили анапсидные черепа, чтобы улучшить свою броню. Более поздние морфологические филогенетические исследования с учётом этого поместили черепах прочно в состав Diapsida. Все молекулярные исследования убедительно подтвердили принадлежность черепах к диапсидам, чаще всего как сестринскую группу по отношению к существующим архозаврам.
Пермские рептилии
С окончанием карбона амниоты стали доминирующими. В то время как примитивные наземные рептилиоморфы всё ещё существовали, синапсидные амниоты представили мегафауну (гигантских животных) в форме пеликозавров, таких как эдафозавр и плотоядный диметродон. В середине пермского периода климат стал более сухим, что привело к изменению фауны: на смену пеликозаврам пришли терапсиды.
Парарептилии, чьи массивные черепа не имели заглазничных отверстий, сохранялись и процветали на протяжении всего пермского периода. Парарептилии из числа парейазавров достигли гигантских размеров в поздней перми и в конце концов исчезли в конце периода (возможно, выжили черепахи).
В начале этого периода современные рептилии развились и разделились на две основные линии: архозавроморфы (предки черепах, крокодилов и динозавров) и лепидозавроморфы (предшественники современных ящериц и туатары). Обе группы оставались ящерицеподобными, относительно небольшими и незаметными в течение пермского периода.
Мезозойские рептилии
В конце пермского периода произошло крупнейшее известное массовое вымирание (см. Пермско-триасовое вымирание). Большая часть более ранней парарептильной и синапсидной мегафауны исчезла, заменившись похожими на современных рептилиями, особенно архозавроморфами. Они характеризовались длинными задними ногами и прямохождением, ранние формы чем-то напоминали длинноногих крокодилов. Архозавры стали доминирующей группой в триасовый период, хотя потребовалось 30 миллионов лет, прежде чем их разнообразие стало таким же большим, как у животных, живших в пермском периоде. Архозавры превратились в хорошо известных нам динозавров и птерозавров, а также предков крокодилов. Поскольку рептилии, сначала рауизухии, а затем динозавры, доминировали в мезозойскую эру, этот интервал широко известен как «эпоха рептилий». Динозавры были как гигантскими, так и мелкими (в том числе более мелких теропод с перьями). В меловой период они дали начало первым настоящим птицам.
Сестринской группой Archosauromorpha является Lepidosauromorpha, в которую входят ящерицы и туатары, а также их ископаемые родственники. Lepidosauromorpha содержал по крайней мере одну основную группу мезозойских морских рептилий: мозазавров, живших в меловой период. Более спорным является филогенетическое размещение других основных групп ископаемых морских рептилий — ихтиоптеригов (включая ихтиозавров) и завроптеригов, которые эволюционировали в раннем триасе. Разные авторы связывали эти группы либо с лепидозавроморфами, либо с архозавроморфами. Итиоптериги также считались диапсидами, которые не принадлежали к наименее инклюзивной кладе, содержащей лепидозавроморфов и архозавроморфов.
Кайнозойские (современные) пресмыкающиеся
В конце мелового периода произошла гибель мегафауны рептилий мезозойской эры (см. Мел-палеогеновое вымирание, знаменитое исчезновением динозавров). Из крупных морских рептилий остались только морские черепахи; а из неморских крупных рептилий только полуводные крокодилы и в целом похожие на них хористодеры пережили вымирание, а последние представители последних, похожие на ящериц Lazarussuchus, вымерли в миоцене. Из огромного множества динозавров, господствовавших в мезозое, выжили только мелкие клювовидные птицы. Эта драматическая модель вымирания в конце мезозоя перешла в кайнозой. Млекопитающие и птицы заполнили пустые ниши, оставленные рептилиями, и, в то время как диверсификация рептилий замедлилась, диверсификация птиц и млекопитающих приняла экспоненциальный оборот. Тем не менее, рептилии по-прежнему были важными организмами фауны.
После исчезновения большинства линий архозавров и морских рептилий к концу мелового периода диверсификация рептилий продолжалась на протяжении всего кайнозоя. Много видов чешуйчатых вымерло во время Мел-палеогенового вымирания, восстановившись только через десять миллионов лет после него. Сегодня чешуйчатые составляют большинство живых рептилий (> 95 %). Известно примерно 11 950 современных видов традиционных рептилий, а птиц ещё около 10 000, что вчетверо превышает количество млекопитающих, представленных примерно 5700 живыми видами.
Классификация
В XIII веке категория рептилий была признана в Европе как состоящая из множества яйцекладущих существ, включая «змей, различных фантастических чудовищ, ящериц, различных амфибий и червей», как записано Винсентом де Бове в его «Зеркале». В XVIII веке рептилии с самого начала классификации были отнесены к амфибиям. Карл Линней, работавший в бедной видами Швеции, где обыкновенная гадюка и ужей уж часто охотятся в воде[прояснить], включил всех рептилий и земноводных в класс «III — Амфибии» в его Systema Naturæ. Термины «рептилия» и «амфибия» были в значительной степени взаимозаменяемы, рептилия (от лат. repere, «ползать») предпочиталась французами. Иосиф Флавий Николаус Лауренти был первым, кто официально использовал термин Reptilia для обозначения рептилий и амфибий, в основном сходного с тем, что у Линнея. Сегодня эти две группы всё ещё изучаются одной наукой герпетологией.
Только в начале XIX века стало ясно, что рептилии и земноводные на самом деле совершенно разные животные, и Пьер Андре Латрейль возвёл для последних класс Baracia (1825 г.), разделив четвероногих на четыре известных класса: рептилий, земноводных, птиц и млекопитающих. Британский анатом Томас Генри Хаксли сделал определение Латрейля популярным и вместе с Ричардом Оуэном расширил Reptilia, включив в него различных ископаемых допотопных монстров, включая динозавров и млекопитающих (синапсид) Dicynodon. Это была не единственная возможная схема классификации: в лекциях Хантера, прочитанных в Королевском колледже хирургов в 1863 году, Хаксли сгруппировал позвоночных в млекопитающих, завроидов и ихтиоидов (последние включали рыб и земноводных). Впоследствии он предложил названия Sauropsida и Ichthyopsida для двух последних групп. В 1866 году Геккель продемонстрировал, что позвоночных можно разделить на основе их репродуктивных стратегий, и что рептилий, птиц и млекопитающих объединяет амниотическое яйцо.
Термины Sauropsida («лица ящериц») и Theropsida («лица зверей») были снова использованы в 1916 году Э. С. Гудричем для различения ящериц, птиц и их родственников, с одной стороны (Sauropsida), и млекопитающих и их вымерших родственников (Theropsida) с другой. Гудрич поддержал это разделение по характеру сердца и сосудов в каждой группе и другим особенностям, таким как строение переднего мозга. По словам Гудрича, обе линии произошли от более ранней основной группы, Protosauria («первые ящерицы»), в которую он включил некоторых животных, которые сегодня считаются рептилиеподобными амфибиями, а также ранних рептилий.
В 1956 году Дэвид Мередит Сирс Уотсон заметил, что первые две группы разошлись очень рано в истории рептилий, поэтому он разделил Protosauria Гудрича между ними. Он также переосмыслил Sauropsida и Theropsida, исключив соответственно птиц и млекопитающих. Таким образом, его Sauropsida включала Procolophonia, Eosuchia, Millerosauria, Chelonia (черепахи), Squamata (ящерицы и змеи), Rhynchocephalia, Crocodilia, «текодонтов» (парафилетические базальные Archosauria), нептичьих динозавров, птерозавров, ихтиозавров и завроптериги.
В конце XIX века был предложен ряд переопределений Рептилий. Признаки, перечисленные Лидеккером в 1896 году, например, включают единственный затылочный мыщелок, челюстной сустав, образованный квадратной и суставной костями, и некоторые характеристики позвонков. Животные, выделяемые этими формулировками, амниоты, отличные от млекопитающих и птиц, до сих пор считаются рептилиями. Разделение синапсид/завропсид дополнило другой подход, который разделил рептилий на четыре подкласса. Эта классификация была инициирована Генри Фэйрфилдом Осборном и развита и стала популярной благодаря классической книге «Палеонтология позвоночных» Ромера. Этими четырьмя подклассами были:
- Anapsida — котилозавры и Chelonia (черепахи и их родственники),
- Synapsida — пеликозавры и терапсиды («звероподобные рептилии»),
- Эвриапсиды — проторозавры (маленькие, ранние ящероподобные рептилии) и морские завроптериги и ихтиозавры, последних в работе Осборна назвали Парапсидами,
- Diapsida — большинство рептилий, включая ящериц, змей, крокодилов, динозавров и птерозавров.
Расположение Euryapsida было неопределённым. Иногда считалось, что ихтиозавры возникли независимо от других эвриапсидов, и им дали более старое название Парапсида. Позже Parapsida была по большей части исключена как группа (ихтиозавры были классифицированы как incertae sedis вместе с Euryapsida). Однако четыре (или три, если Euryapsida сливаются с Diapsida) подкласса оставались более или менее универсальными для неспециалистов на протяжении всего XX века. Недавние исследователи в значительной степени отказались от него.
К началу XXI века палеонтологи позвоночных начали принимать филогенетическую таксономию, в которой все группы определяются таким образом, чтобы быть монофилетическими; то есть группы, в которые входят все потомки определённого предка. Рептилии, как исторически определено, являются парафилетическими, поскольку они исключают как птиц, так и млекопитающих. Они соответственно произошли от динозавров и от ранних терапсидов, которых традиционно называли рептилиями. Птицы более тесно связаны с крокодилами, чем последние с остальными современными рептилиями. Несмотря на ранние предложения о замене парафилетических рептилий монофилетическими завропсидами, включающими птиц, этот термин так и не получил широкого распространения, а если и применялся, то не применялся последовательно.
Колин Тадж писал:
Млекопитающие являются кладой, и поэтому кладисты рады признать традиционный таксон млекопитающих; и птицы тоже представляют собой кладу, повсеместно приписываемую формальному таксону Aves. Mammalia и Aves, по сути, являются субкладами в большой кладе Amniota. Но традиционный класс Reptilia не является кладой. Это всего лишь часть клады Amniota: часть, оставшаяся после отделения Mammalia и Aves. Он не может быть определён синапоморфно, как это правильно. Вместо этого он определяется комбинацией свойств, которые у него есть, и признаков, которых ему не хватает: рептилии — это амниоты, у которых нет меха или перьев. В лучшем случае, предполагают кладисты, мы могли бы сказать, что традиционные рептилии — это «амниоты, не относящиеся к птицам и млекопитающим».
Оригинальный текст (англ.)[показатьскрыть]Mammals are a clade, and therefore the cladists are happy to acknowledge the traditional taxon Mammalia; and birds, too, are a clade, universally ascribed to the formal taxon Aves. Mammalia and Aves are, in fact, subclades within the grand clade of the Amniota. But the traditional class Reptilia is not a clade. It is just a section of the clade Amniota: the section that is left after the Mammalia and Aves have been hived off. It cannot be defined by synapomorphies, as is the proper way. Instead, it is defined by a combination of the features it has and the features it lacks: reptiles are the amniotes that lack fur or feathers. At best, the cladists suggest, we could say that the traditional Reptilia are 'non-avian, non-mammalian amniotes'.
Когда использовался термин Sauropsida, она имел почти то же определение, что и Reptilia. В 1988 году Жак Готье предложил кладистическое определение Reptilia как монофилетической группы, включающей черепах, ящериц и змей, крокодилов и птиц, их общего предка и всех его потомков. Хотя определение Готье было близко к современному консенсусу, тем не менее, оно стало считаться неверным, потому что фактическая связь черепах с другими рептилиями в то время ещё не была хорошо изучена. С тех пор основные изменения включали переназначение синапсид как нерептилий и классификацию черепах как диапсид.
Множество других определений было предложено другими учёными в годы после статьи Готье. Первое такое новое определение, было опубликовано Модесто и Андерсоном в 2004 году. Они определили Reptilia как всех амниот, более близких к Lacerta agilis и Crocodylus niloticus, чем к Homo sapiens. Это основанное на стебле определение эквивалентно более распространённому определению Sauropsida, которое Модесто и Андерсон синонимизировали с Reptilia, поскольку последнее более известно и чаще используется. Однако, в отличие от большинства предыдущих определений Reptilia, определение Модесто и Андерсона включает птиц, поскольку они входят в кладу, включающую как ящериц, так и крокодилов.
Дочерние таксоны



По часовой стрелке, начиная сверху слева: гаттерия, чёрная мамба, обыкновенная игуана, Smaug breyeri и сетчатый питон.

Сверху: краснобрюхая короткошейная черепаха, индийская лопастная черепаха;
Снизу: галапагосская черепаха, бисса.

1-й ряд: Erythrosuchus africanus, эвпаркерия;
2-й ряд: филиппинский крокодил, Typothorax coccinarum;
3-й ряд: шлемоносный казуар, Anhanguera piscator.
Класс Пресмыкающиеся (Reptilia)
-
† Подкласс Парарептилии (Parareptilia) — возможно, базальные диапсиды
- † Отряд Мезозавры (Mesosauria)
- † Отряд Millerosauria
- † Отряд Проколофономорфы (Procolophonomorpha)
- † ? Отряд Болозавры (Bolosauria) — возможно, в отряде Procolophonomorpha
-
Подкласс Диапсиды (Diapsida)
- Отряд † Ареосцелидии (Araeoscelidia)
- Клада Неодиапсиды (Neodiapsida)
- Отряд † Талаттозавры (Thalattosauria)
- Клада † Drepanosauromorpha
- Клада † Ichthyosauromorpha — положение спорно
- Надотряд † Ихтиоптеригии (Ichthyopterygia)
- Отряд † Ихтиозавры (Ichthyosauria)
- Надотряд † Ихтиоптеригии (Ichthyopterygia)
-
Клада Заврии (Sauria)
-
incertae sedis
- Отряд † Хористодеры (Choristodera) — возможно, базальные архозавроморфы
- Надотряд † Завроптеригии (Sauropterygia) — возможно, парафилетическая группа к Testudinata
- Отряд † Плакодонты (Placodontia)
- Отряд † Nothosauroidea
- Отряд † Плезиозавры (Plesiosauria)
-
Инфракласс Лепидозавроморфы (Lepidosauromorpha)
- Надотряд Лепидозавры (Lepidosauria)
- Отряд Клювоголовые (Rhynchocephalia)
- Отряд Чешуйчатые (Squamata)
- Надотряд Лепидозавры (Lepidosauria)
-
Инфракласс Архозавроморфы (Archosauromorpha s. l.) [ ≈ Archelosauria]
- Клада Тестудинаты (Testudinata)
- Отряд Черепахи (Testudines)
- Клада Архозавроморфы (Archosauromorpha s. s.)
- Клада Crocopoda
- Клада † Allokotosauria
- Отряд † Ринхозавры (Rhynchosauria)
- Клада Архозавриформы (Archosauriformes)
- Отряд † Фитозавры (Phytosauria) и более базальные таксоны
- Клада Архозавры (Archosauria)
- Клада Псевдозухии (Pseudosuchia)
- Отряд † Этозавры (Aetosauria) и прочие базальные таксоны
- Надотряд Крокодиломорфы (Crocodylomorpha)
- Отряд Крокодилы (Crocodilia)
- Клада Авеметатарзалии (Avemetatarsalia)
- Клада † Aphanosauria
- Клада Орнитодиры (Ornithodira)
- Отряд † Птерозавры (Pterosauria)
- Надотряд Динозавры (Dinosauria)
- Отряд Ящеротазовые (Saurischia) — в кладистике включает птиц (Aves)
- Отряд † Птицетазовые (Ornithischia)
- Клада Псевдозухии (Pseudosuchia)
- Клада Crocopoda
- Клада Тестудинаты (Testudinata)
-
incertae sedis
Филогенетика
Согласно определению Modesto & Anderson, 2004, пресмыкающиеся, или рептилии (Reptilia), являются наиболее инклюзивной кладой, включающей Lacerta agilis Linnaeus, 1758 и Crocodylus niloticus Laurenti, 1768, но не Homo sapiens Linnaeus, 1758. Данное определение эквивалентно определению завропсид (Sauropsida), данному Готье в 1994 году, согласно которому к этой кладе относятся все пресмыкающиеся и другие амниоты, ближе родственные им, чем млекопитающим (Mammalia).
Кладограмма согласно Lee, 2013:
| Амниоты |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||





Кладограмма, основанная на Ford & Benson, 2020:
| Амниоты |
|
||||||||||||||||||||||||||||||||||||||||||


Строение
Пресмыкающиеся, наряду с птицами и млекопитающими относятся к высшим позвоночным животным — амниотам, и в целом их строение типично для данной группы, однако, также имеются черты более простых по строению амфибий.
Покров



Наружный кожный покров современных пресмыкающихся в результате утолщения и ороговения образует чешуйки или щитки. У ящериц роговые чешуйки перекрывают друг друга, напоминая черепицу. У черепах сросшиеся щитки формируют сплошной прочный панцирь. Смена рогового покрова происходит путём полной или частичной линьки, которая у многих видов происходит несколько раз в год.
Наряду с этим, древние рептилии могли также быть покрыты своеобразными перьями (являющимися по строению специализированной формой чешуи) или похожими на пух нитевидными образованиями (прогрессивные архозавры).
Плотная и сухая кожа содержит пахучие железы. Слизистые железы отсутствуют — кожа рептилий сухая, характерный блеск ей придают гладкие чешуи.
В наружной части внутреннего слоя кожи часто находятся специальные клетки — хроматофоры. В этих клетках секретируются пигменты: меланины и каротиноиды. Также в хроматофорах встречается способный отражать свет гуанин. Благодаря хроматофорам, некоторые пресмыкающиеся способны изменять окраску своего тела за сравнительно короткое время. Хамелеоны — наиболее известные представители с подобным свойством.
Скелет
В осевом скелете пресмыкающихся разделение на отделы более заметно, чем у земноводных. Хорошо различимы пять отделов позвоночника: шейный (лат. pars cervicalis), туловищный (пояснично-грудной, pars thoracolumbalis), поясничный, крестцовый (pars sacralis) и хвостовой (pars caudalis).
Типичным для пресмыкающихся является следующее строение осевого скелета. Общее количество позвонков различно у разных видов (50—80, у змей возрастает до 140—435). Из позвонков шейного отдела (от 7 до 10) два передних (атлант и эпистрофей) образуют сустав, позволяющий голове не только двигаться в вертикальной плоскости относительно первого шейного позвонка, но и поворачиваться. В туловищном отделе от 16 до 25 позвонков, каждый с парой рёбер. Первые несколько пар рёбер прикрепляются к грудине, образуя грудную клетку (отсутствует у змей). В крестцовом отделе всего два позвонка, к широким поперечным отросткам которых причленяется таз. Хвостовой отдел составляют несколько десятков (15—40) постепенно уменьшающихся в размерах позвонков. Последние хвостовые позвонки представляют собой небольшие палочковидные косточки.
В некоторых группах рептилий осевой скелет имеет отличия. У змей позвоночник отчётливо делится лишь на туловищный и хвостовой отделы, грудина отсутствует. У черепах позвонки туловищного отдела срастаются со спинным щитом панциря, вследствие чего неподвижны.
Череп пресмыкающихся значительно более окостеневший, чем у земноводных. Лишь в обонятельной капсуле и слуховой области содержится небольшое количество хряща. Осевой и висцеральный отделы черепа эмбрионально формируются отдельно, но у взрослых особей срастаются в единое образование. В состав черепа входят как хрящевые (замещающие, или первичные), так и многочисленные кожные (покровные, или вторичные) кости.
Пояс передних конечностей сходен с поясом земноводных, отличаясь лишь более сильным развитием окостенения. Пара передних конечностей рептилий состоит из плеча, предплечья и кисти. Пара задних конечностей — из бедра, голени и стопы. На фалангах конечностей расположены когти.
Мышечная система
Мышечная система пресмыкающихся представлена жевательной, шейной мускулатурой, мускулатурой брюшного пресса, а также мускулатурой сгибателей и разгибателей. Присутствуют характерные для амниот межрёберные мышцы, играющие важную роль при акте дыхания. Подкожная мускулатура позволяет изменять положение роговых чешуй.
Энергоснабжение произвольной мускулатуры современных пресмыкающихся происходит главным образом за счёт анаэробного разложения глюкозы — порядка 50—75 % мышечных волокон в их мышцах приспособлены к работе в анаэробных условиях (без участия кислорода). Такой метаболизм позволяет им перемещаться на короткие расстояния столь же быстро, как и теплокровным животным, и обеспечивает даже большую силу сокращения мышц, причём интенсивность реакции у многих групп слабо зависит от температуры тела. Однако уже через 1—2 минуты интенсивной работы мускулатуры в анаэробном режиме в ней накапливается молочная кислота, что приводит к наступлению своеобразной мышечной усталости и изменению pH крови. В результате уже после нескольких быстрых пробежек рептилия становится практически неспособна к активным действиям на время от нескольких минут до нескольких часов, требующееся для распада накопившейся в мышцах молочной кислоты. Более крупные животные способны дольше терпеть накопление в мышцах молочной кислоты и таким образом фактически являются более выносливыми. Крокодилы способны выдерживать падение pH крови до 6,42 и ниже. Некоторые современные рептилии, в частности, вараны и кожистая черепаха, тем не менее, являются исключением из этого правила: в ходе активности их уровень метаболизма может достигать более половины от такового у теплокровных животных сопоставимой массы, и они в значительной мере полагаются на аэробный метаболизм, почти не испытывая проблем с накоплением молочной кислоты.
Среди вымерших древних рептилий известны высокоподвижные формы (такие как архозавры: наземные крокодиломорфы, динозавры и птерозавры, а также лепидозавры: мозазавры, ихтиозавры и завроптеригии), строение тела и предполагаемый образ жизни которых говорят о способности поддерживать продолжительную мышечную активность без утомления, что соответствует более интенсивному, чем у современных пресмыкающихся, обмену веществ, и работе мышц преимущественно по аэробному механизму, требующему интенсивного снабжения кислородом. Это в свою очередь должно было сопровождаться совершенно другой физиологией организма (полное разделение желудочков сердца, повышение эффективности дыхания за счёт отделения воздухоносных путей от ротовой полости, появление эффективных механизмов терморегуляции и так далее). Данные изменения отчасти прослеживаются на палеонтологическом материале в линии рептилий, ведущей к птицам. В настоящее время надёжно показано существование теплокровности как минимум у некоторых групп динозавров и предков современных крокодилов, а также плезиозавров, мозазавров и ихтиозавров.
Нервная система

Как и у большинства хордовых животных, центральная нервная система пресмыкающихся представлена головным (из 5 отделов) и спинным мозгом.
Головной мозг расположен внутри черепа. Ряд важных особенностей отличает головной мозг пресмыкающихся от головного мозга земноводных. Нередко говорят[кто?] о так называемом завропсидном типе головного мозга, присущем также и птицам, в отличие от ихтиопсидного типа у рыб и земноводных.
Выделяют пять отделов головного мозга пресмыкающихся.
- Передний мозг состоит из двух больших полушарий, от которых отходят обонятельные доли. Поверхность больших полушарий абсолютно гладкая. В мозговом своде полушарий различают первичный свод — архипаллиум, занимающий большую часть крыши полушарий, и зачатки неопаллиума. Дно переднего мозга в основном состоит из полосатых тел.
- Промежуточный мозг расположен между передним и средним мозгом. В верхней его части расположен эпифиз, а на нижней стороне — гипофиз. У большинства ящериц и гаттерии (а также у многих вымерших форм) рядом с эпифизом развивается теменной глаз, а у крокодилов потеряны оба этих органа. Дно промежуточного мозга занято зрительными нервами и их перекрестом (хиазмой).
- Средний мозг представлен двумя крупными передними холмами — зрительными долями, а также небольшими задними холмами. Зрительная кора более развита, чем у амфибий.
- Мозжечок прикрывает переднюю часть продолговатого мозга. Он более крупный по сравнению с мозжечком амфибий.
- Продолговатый мозг образует изгиб в вертикальной плоскости, который характерен для всех амниот.
От головного мозга отходят 12 пар черепных нервов. В спинном мозге разделение на белое и серое вещество более отчётливо, чем у амфибий. От спинного мозга отходят сегментальные спинномозговые нервы, образуя типичное плечевое и тазовое сплетение. Отчётливо выражена вегетативная нервная система (симпатическая и парасимпатическая) в виде цепи парных нервных ганглиев.
Органы чувств

Пресмыкающиеся имеют шесть основных органов чувств, из которых зрение имеет решающее значение.
- Орган зрения — глаза, устроены сложнее, чем у амфибий: в склере присутствует кольцо из тонких костных пластинок; от задней стенки глазного яблока отходит вырост — гребешок, вдающийся в стекловидное тело; в ресничном теле развита поперечно-полосатая мускулатура, которая позволяет не только перемещать хрусталик, но и изменять его форму, таким образом осуществляя наводку на резкость в процессе аккомодации. Органы зрения имеют приспособления к работе в воздушной среде. Слёзные железы предохраняют глаза от высыхания. Наружные веки и мигательная перепонка выполняют защитную функцию. У змей и некоторых ящериц веки срастаются, формируя прозрачную оболочку. Сетчатка глаза может содержать как палочки, так и колбочки. У ночных видов колбочки отсутствуют. У большинства дневных видов диапазон цветного зрения смещён в жёлто-оранжевую часть спектра. Форма зрачка чаще всего круглая или в виде вертикальной щели (как у кошки), горизонтальный щелевидный зрачок, весьма распространённый у земноводных, среди рептилий встречается очень редко, например у плетевидных змей. Теменной глаз, сохранившийся у некоторых видов, важен для ориентирования в пространстве и синхронизации суточных ритмов организма с циклом смены дня и ночи, хотя многое в его функциях ещё неясно.
- Орган обоняния представлен внутренними ноздрями — хоанами и вомероназальным органом. По сравнению со строением земноводных, хоаны расположены ближе к глотке, что даёт возможность свободно дышать в то время, как пища находится во рту. Обоняние развито лучше, чем у земноводных, позволяя многим ящерицам находить пищу, находящуюся под поверхностью песка на глубине до 6—8 см.
- Орган вкуса — вкусовые луковицы, расположенные в основном в глотке.
- Орган тепловой чувствительности находится на лицевой ямке между глазом и носом с каждой стороны головы. Особенно развит у змей. У ямкоголовых змей термолокаторы позволяют определять даже направление источника теплового излучения.
- Орган слуха близок к органу слуха лягушек, он содержит внутреннее и среднее ухо, снабжённое барабанной перепонкой, слуховой косточкой — стременем и евстахиевой трубой. Роль слуха в жизни пресмыкающихся сравнительно невелика, особенно слаб слух у змей, не имеющих барабанной перепонки и воспринимающих колебания, распространяющиеся по земле или в воде. Пресмыкающиеся воспринимают звуки в диапазоне 20—6000 Гц, хотя большинство хорошо слышит лишь в диапазоне 60—200 Гц (у крокодилов 100—3000 Гц).
- Осязание выражено отчётливо, особенно у черепах, которые могут чувствовать даже лёгкое прикосновение к панцирю.
Дыхательная система
Для пресмыкающихся характерно дыхание всасывающего типа путём расширения и сужения грудной клетки при помощи межрёберной и брюшной мускулатуры. Попавший через гортань воздух поступает в трахею — длинную дыхательную трубку, которая на конце делится на бронхи, ведущие в лёгкие. Как и у земноводных, лёгкие пресмыкающихся имеют мешкообразное строение, хотя их внутренняя структура намного сложнее. Внутренние стенки лёгочных мешков имеют складчатое ячеистое строение, что значительно увеличивает дыхательную поверхность. Строение лёгких значительно отличается у разных видов и может представлять слабодифференциированные органы с малым количеством ячей, у примитивных видов, таких как гаттерии, среднеразвитые органы с ярко выраженной внутренней структурой, как у большинства чешуйчатых, так и высокоразвитые лёгкие, имеющие почти губчатое строение (как у птиц и млекопитающих), у таких рептилий, как вараны, тегу, крокодилы, многие черепахи и крупные змеи. Водные черепахи имеют более проницаемую кожу, а некоторые виды даже изменили свою клоаку, чтобы увеличить площадь газообмена. Даже с этими адаптациями дыхание никогда полностью не осуществляется без лёгких. Смена воздуха в лёгких у каждой основной группы рептилий осуществляется по-разному. У чешуйчатых лёгкие вентилируются почти исключительно осевой мускулатурой. Это та же самая мускулатура, которая используется при передвижении. Из-за этого ограничения большинство чешуек вынуждены задерживать дыхание во время интенсивных пробежек. Некоторые, однако, нашли способ обойти это. Вараниды и некоторые другие виды ящериц используют буккальное сцеживание как дополнение к их обычному «аксиальному дыханию». Это позволяет животным полностью наполнять свои лёгкие во время бега и, таким образом, длительное время оставаться аэробно активными. Известно, что у ящериц тегу имеется протодиафрагма, отделяющая лёгочную полость от висцеральной. Хотя на самом деле он не способен к движению, он позволяет сильнее раздувать лёгкие, снимая вес внутренних органов с них.
Поскольку тело покрыто чешуйками, кожное дыхание у пресмыкающихся отсутствует (исключение составляют мягкотелые черепахи и морские змеи), и лёгкие являются единственным дыхательным органом.
У видов, живущих в воде, могут появляться специализированные адаптации, для того чтобы обходиться дольше без атмосферного воздуха, например у морских змей это слизистая выстилка рта, а у черепах это и полость рта и специальные анальные мешки, способные усваивать кислород из воды.
Крокодилы на самом деле имеют мышечную диафрагму, аналогичную диафрагме млекопитающих. Разница в том, что мышцы крокодиловой диафрагмы тянут лобковую кость (часть таза, подвижную у крокодилов) назад, что опускает печень, тем самым освобождая пространство для расширения лёгких. Этот тип диафрагмальной установки называют «печёночным поршнем». Дыхательные пути образуют ряд двойных трубчатых камер в каждом лёгком. При вдохе и выдохе воздух проходит через дыхательные пути в одном направлении, создавая таким образом однонаправленный поток воздуха через лёгкие. Аналогичная система обнаружена у птиц, варанов и игуан.
У большинства рептилий нет вторичного неба, а это означает, что они должны задерживать дыхание при глотании. Крокодилы развили костлявое вторичное нёбо, которое позволяет им продолжать дышать, оставаясь под водой (и защищать свой мозг от повреждений, связанных с борьбой с добычей). Сцинки (семейство Scincidae) также в той или иной степени развили костное вторичное небо. Змеи использовали другой подход и вместо этого расширили свои трахеи. Их расширение трахеи торчит изо рта в виде трубки, и позволяет этим животным проглатывать крупную добычу, не страдая от удушья.
Система кровообращения
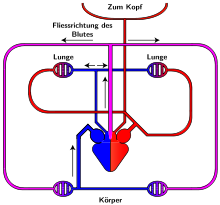
Как и амфибии, большинство пресмыкающихся обладают трёхкамерным сердцем, состоящим из желудочка и двух предсердий. Желудочек разделён неполной перегородкой на две половины: верхнюю и нижнюю, и может быть условно разделён на три отдела. Однако трёхкамерное сердце рептилий развилось независимо от такового у земноводных, и различается в плане строения (и иногда — функциональности) у разных современных групп.
Все лепидозавры и черепахи имеют трёхкамерное сердце, состоящее из двух предсердий, одного желудочка с переменными перегородками и двух аорт, ведущих в большой круг кровообращения. Степень смешения оксигенированной и деоксигенированной крови в трёхкамерном сердце различна в зависимости от вида и физиологического состояния. При различных условиях деоксигенированная кровь может быть отправлена обратно в организм, а насыщенная кислородом кровь может быть отправлена обратно в лёгкие. Было высказано предположение, что это изменение кровотока обеспечивает более эффективную терморегуляцию и более длительное время погружения для водных видов, но не было доказано, что оно является преимуществом и для наземных видов.
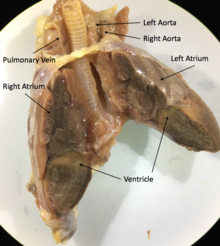
Будучи архозаврами, крокодилы обладают четырёхкамерным сердцем. Перегородка полностью разделяет желудочек на две половины: правую — венозную и левую — артериальную, таким образом обосабливая полноценное четырёхкамерное сердце, как у млекопитающих и птиц. Лишь после образования полных перегородок у крокодилов образуется отверстие Панница в ходе эмбриогенеза, допускающее контролируемое смешивание артериальной и венозной крови, что связано со вторичным понижением уровня обмена веществ у этой группы животных. Крокодилы имеют анатомически четырёхкамерное сердце, похожее на птичье, но также имеют две аорты.
Например, сердце игуаны, как и большинство чешуйчатых сердец, состоит из трёх камер с двумя аортами и одним желудочком, сердечными непроизвольными мышцами. Основными структурами сердца являются венозный синус, водитель ритма, левое предсердие, правое предсердие, атриовентрикулярный клапан, венозная полость, артериальная полость, лёгочная полость, мышечный гребень, вентрикулярный гребень, лёгочные вены и парные дуги аорты.
Некоторые виды чешуйчатых (например, питоны и вараны) имеют трёхкамерное сердце, которое при сокращении функционально становится четырёхкамерным. Это стало возможным благодаря мышечному гребню, который разделяет желудочек во время диастолы желудочков и полностью разделяет его во время систолы желудочков . Из-за этого гребня некоторые из этих чешуек способны создавать перепады давления в желудочках, эквивалентные тем, которые наблюдаются в сердцах млекопитающих и птиц.
Полного разделения на два независимых круга кровообращения у пресмыкающихся не происходит, поскольку венозная и артериальная кровь смешиваются в спинной аорте.
Подобно рыбам и амфибиям, подавляющее большинство современных пресмыкающихся — холоднокровные животные. Несмотря на это, многие пресмыкающиеся могут регулировать температуру, перемещаясь из тени на солнце и обратно или меняя цвет, становясь темнее, чтобы согреться, или светлее, чтобы остыть. Сравнительно недавно у ящериц тегу была обнаружена факультативная эндотермия — возможность повышать температуру тела во время брачного периода за счёт эндотермических химических реакций. Вараны в разгар пищевой активности также могут повышать температуру тела за счёт внутренних процессов. Кожистая черепаха, в свою очередь, имеет изолирующую жировую прослойку и за счёт интенсивной мышечной активности может быть примерно на 18 градусов теплее окружающей её воды.
Метаболизм
Современные нептичьи рептилии холоднокровны, так что они имеют ограниченные физиологические средства поддержания постоянной температуры тела и часто полагаются на внешние источники тепла. Из-за менее стабильной внутренней температуры, чем у птиц и млекопитающих, биохимия рептилий требует ферментов, способных сохранять эффективность в более широком диапазоне температур, чем в случае теплокровных .животные. Оптимальный диапазон температуры тела зависит от вида, но обычно ниже, чем у теплокровных животных; для многих ящериц она находится в диапазоне 24-35 °C (75-95 °F), в то время как у экстремально адаптированных к жаре видов, таких как американская пустынная игуана Dipsosaurus dorsalis, могут быть оптимальные физиологические температуры для млекопитающих, в диапазоне от 35° до 40 °C (от 95° до 104 °F). В то время как оптимальная температура часто наблюдается, когда животное активно, низкий основной обмен приводит к быстрому падению температуры тела, когда животное неактивно.
Как и у всех животных, мышечная деятельность рептилий производит тепло. У крупных рептилий, таких как кожистые черепахи, низкое отношение поверхности к объёму позволяет этому метаболически вырабатываемому теплу держать животных теплее, чем их окружающая среда, даже если у них нет теплокровного метаболизма. Эта форма гомеотермии называется гигантотермией; было высказано предположение, что она была обычным явлением у крупных динозавров и других вымерших крупных рептилий.
Преимущество низкого метаболизма в состоянии покоя заключается в том, что для поддержания функций организма требуется гораздо меньше пищи. Используя колебания температуры окружающей среды или оставаясь холодными, когда им не нужно двигаться, рептилии могут экономить значительное количество энергии по сравнению с эндотермическими животными того же размера. Крокодилу требуется от одной десятой до одной пятой части пищи, необходимой льву того же веса, и он может прожить без еды полгода. Более низкие потребности в пище и адаптивный метаболизм позволяют рептилиям доминировать в животной жизни в регионах, где доступность пищи слишком мала для поддержания крупных млекопитающих и птиц.
Принято считать, что рептилии не могут производить устойчиво высокий уровень энергии, необходимый для погони на большие расстояния или полёта. Более высокая энергетическая ёмкость могла быть ответственна за эволюцию теплокровности у птиц и млекопитающих. Однако исследование корреляции между активной ёмкостью и термофизиологией показывает слабую связь. Большинство современных рептилий — плотоядные животные со стратегией кормления «сиди и жди»; неясно, являются ли рептилии хладнокровными из-за их экологии. Энергетические исследования некоторых рептилий показали, что их активные способности равны или превосходят возможности теплокровных животных аналогичного размера.
Пищеварительная система
Вследствие разнообразия доступной пищи[прояснить]пищеварительный тракт у пресмыкающихся гораздо более дифференцирован, чем у земноводных животных.
Пищеварительная система начинается ротовым отверстием, ограниченным челюстями с обычно коническими, одинаковыми, прочно прирастающими зубами (гомодонтная система, хотя у некоторых вымерших форм была гетеродонтная). У рептилий, за исключением крокодилов и большинства черепах, есть гибкие неподвижные губы, плотно закрывающие рот и зубы в состоянии покоя. Зубы пресмыкающихся не предназначены для жевания — они служат для захвата и удержания пищи, реже — для её разделения или отрывания удобных по размеру кусков, у видов, питающихся моллюсками и насекомыми, зубы приспособлены для дробления жёстких покровов этих животных. Пища практически всегда проглатывается целиком, как животная, так и растительная. Язык свободный, впереди мускулистый, подвижный, к концу утончается и раздваивается. У некоторых видов язык клейкий, и служит для ловли добычи. Язык служит для проглатывания пищи, но виды с тонким и длинным языком проталкивают пищу в глотку толчками головы (ящерицы) или движением челюстей (змеи) Ротовая полость отграничена от глотки развивающимся вторичным костным нёбом у ящериц и черепах, полностью развитым костным небом у крокодилов, но у змей и варанов вторичное костное небо отсутствует. Многоклеточные слюнные железы содержат пищеварительные ферменты. Глотка переходит в узкий, но очень растяжимый пищевод, далее в мускулистый желудок и кишечник. Желудок имеет толстые мышечные стенки. На границе между тонкой и толстой кишкой находится слепая кишка, которой нет у амфибий. Большая печень рептилий имеет жёлчный пузырь. Поджелудочная железа в виде длинного плотного тела лежит в петле двенадцатиперстной кишки. Кишечник заканчивается клоакой.
Выделительная система

Выделительная система пресмыкающихся представлена почками, мочеточниками и мочевым пузырём.
Почки пресмыкающихся существенно отличаются от почек рыб и амфибий, которым приходится решать задачу избавления от постоянного избытка воды в организме. Вместо туловищных почек амфибий (мезонефрос), почки рептилий (метанефрос) располагаются в тазовой области с брюшной стороны клоаки и по её бокам. Почки соединяются с клоакой через мочеточники.
Тонкостенный стебельчатый мочевой пузырь соединяется с клоакой тонкой шейкой на её брюшной стороне. У некоторых пресмыкающихся мочевой пузырь недоразвит (крокодилы, змеи, некоторые ящерицы).
У наземных рептилий в качестве конечного продукта азотистого обмена выступает слабо растворимая в воде мочевая кислота, выделяемая вместе с калом через клоаку, что позволяет более эффективно сохранять влагу.
Половая система
Пресмыкающиеся — раздельнополые животные, двуполое размножение.
Половая система самцов состоит из пары семенников, которые расположены по бокам поясничного отдела позвоночника. От каждого семенника отходит семенной канал, который впадает в вольфов канал. С появлением тазовой почки у пресмыкающихся вольфов канал у самцов выступает лишь как семяпровод и полностью отсутствует у самок. Вольфов канал открывается в клоаку, образуя семенной пузырёк.
Половая система самок представлена яичниками, которые подвешены на брыжейке к спинной стороне полости тела по бокам позвоночника. Яйцеводы (мюллеровы каналы) также подвешены на брыжейке. В переднюю часть полости тела яйцеводы открываются щелевидными отверстиями — воронками. Нижний конец яйцеводов открывается в нижний отдел клоаки на её спинной стороне.
Образ жизни
Развитие
Оплодотворение внутреннее. Развитие зародыша происходит в яйце с кожистой или известковой оболочкой, наряду с этим встречается яйцеживорождение и (реже) истинное живорождение. У пресмыкающихся прямое постэмбриональное развитие. Для многих представителей характерна забота о потомстве, в частности самки крокодилов переносят потомство от места кладки к водоёмам в ротовой полости, хотя в некоторых случаях могут съесть детёныша.
Питание
Большинство пресмыкающихся относится к плотоядным животным (включая насекомоядность и поедание падали, истинное хищничество среди современных рептилий свойственно только крокодилам, ряду ящериц и большинству змей). Для некоторых (например, агамы, игуаны) характерно смешанное питание. Иногда встречаются и почти исключительно травоядные пресмыкающиеся (сухопутные черепахи).
Передвижение


Примитивные группы древних амниот (в том числе первые рептилии) изначально передвигались на четырёх широко расставленных в стороны конечностях, которые при локомоции раздвигались медиалатерально и не были способны постоянно поддерживать туловище над землёй, отсюда — «пресмыкание», волочение живота по земле, давшее русскоязычное название всей этой группе, что соответствовало характерному для них низкому уровню интенсивности обмена веществ и сравнительно малоподвижному образу жизни, связанному с чередованием коротких вспышек активности и длительного отдыха лёжа на животе. Современные пресмыкающиеся в основном сохраняют подобный, базальный в эволюционном плане, характер локомоции, некоторое исключение составляют только крокодилы, у которых конечности при беге могут занимать положение почти под туловищем. Причём у некоторых вымерших групп современного отряда крокодилов, которые вели исключительно сухопутный образ жизни, такое положение конечностей было габитуальным, то есть поддерживалось большую часть времени — в Австралии такие наземные крокодилы (например, Квинкана) обитали ещё 40 тыс. лет назад, то есть одновременно с человеком современного вида. У варанов конечности полувыпрямлены, что связано с активным перемещением, и при беге туловище находится высоко над землёй. Змеи и безногие ящерицы так же является исключением, так как отсутствие конечностей и совершенно иной способ локомоции (ползание), чем у рептилий с конечностями, обеспечивают быстрое перемещение большинству змей по любой поверхности.
Большинство рептилий неплохо плавают за счёт «рыбообразного» изгибания тела в поперечной плоскости. У некоторых групп приспособление к водной среде достигло более высокой степени, причём в процессе эволюции у них вырабатывались различные типы локомоции. Так, ихтиозавры, мозазавры и, в меньшей степени, крокодилы сохранили характер плавания за счёт поперечных изгибов туловища. С другой стороны, у морских черепах и вымерших плезиозавров выработался новый механизм движения в воде, за счёт взмахов конечностей, преобразовавшихся в ласты. Многие эти группы практически полностью утратили способность перемещаться на суше. Несколько родов пресмыкающихся способны к планирующему полёту — например летучие драконы (агамы рода Draco) и украшенные древесные змеи. Активно летающие рептилии известны только из ископаемых остатков (см. Птерозавры).
В древности развитие некоторых продвинутых групп рептилий шло в иных направлениях — их конечности стали перемещаться непосредственно под туловище, занимая более габитуально вертикальное (выпрямленное) положение, что соответствовало более подвижному образу жизни, связанному с быстрым бегом и активным преследованием добычи.

У предков архозавров лобковая и седалищная кости начали удлиняться, передние ноги — укорачиваться, а тяжёлый длинный хвост, практически лишившийся подвижности из-за срастания тазовых костей, превратился в балансир, уравновешивающий массу вытянутого туловища. Задние конечности при ходьбе стали перемещаться в парасагиттальной плоскости, голеностопный сустав получил возможность пружинить при ходьбе, что способствовало более экономному расходу сил, а опора переместилась на средний (третий) палец. Это позволило многим группам текодонтов перейти к факультативной, или даже габитуальной, бипедальности, подобно современным бегающим птицам, специализировавшись на беге на двух ногах.
Данные изменения должны были сопровождаться значительным повышением интенсивности обмена веществ, поскольку постоянное поддержание тела над землёй и длительная локомоторная активность требовали намного больших энергозатрат по сравнению с «пресмыканием». Однако проследить подобные изменения на палеонтологическом материале сложно. Достоверно известно, однако, что потомки данной группы рептилий — птицы — имеют интенсивный обмен веществ и эндотермную систему терморегуляции (теплокровность).
Ранние динозавры, как ящеротазовые, так и птицетазовые — были активными хищниками, как минимум часть времени передвигавшимися на задних конечностях, что способствовало их быстрому прогрессу, особенно после вымирания большинства синапсидов на рубеже пермского и триасового периода. В юрском периоде почти все более-менее крупные наземные животные (свыше метра в длину) были динозаврами.
Постепенно часть динозавров переходит на растительную пищу, занимая освободившиеся экологические ниши. Большинство растительноядных видов вновь возвращается к передвижению на четырёх конечностях, эволюция же хищников продолжает идти по «двуногому» пути (например, у тираннозаврид и абелизаврид передние конечности оказались практически редуцированны, а у манирапторов — напротив, развились в специализированную хватательную форму). В течение 140 млн лет, вплоть до конца мелового периода, основными наземными хищными позвоночными были двуногие динозавры, некоторые формы которых по большинству признаков неотличимы от птиц.
Как видно, исторически для рептилий было свойственно главным образом именно активное передвижение на двух или четырёх выпрямленных конечностях, именно такие бегающие формы занимали господствующее положение в данном классе (парафилетической группе) животных на протяжении большей части его истории. Двуногие хищные формы изредка возникали у птиц — например фороракосы. В настоящее время подобные формы среди рептилий не возникают, поскольку экологические ниши, связанные с активным преследованием добычи, ныне заняты млекопитающими, и организм современных рептилий требует значительной перестройки для их освоения. Некоторое исключение составляет насекомоядная плащеносная ящерица, и некоторые другие агамовые, способные к бегу на двух ногах (при сохранении расположения конечностей по бокам от туловища). Такая особенность позволяет им обходить ограничение Карриера (невозможность бежать на четырёх ногах и дышать одновременно) и активно преследовать добычу или спасаться от хищников. Ящерица шлемоносный василиск во время бипедальной локомоции способна также бегать по поверхности воды. Тегу и вараны также научились обходить ограничение Карриера, но за счёт модификации дыхательной системы, а не подъёма на задние конечности.
Издаваемые звуки
Большинство рептилий не имеет настоящего голосового аппарата и производят лишь самые примитивные звуки вроде шипения или свиста. Крокодилы рычат, издают лающие и рокочущие звуки, детёныши квакают. Черепахи в некоторых случаях (испуг, брачный сезон, боль) могут издавать писк или стон. Представители гекконовых ящериц издают самые разные звуки — писк, визг, щебет, щелчки, крики.
Хозяйственное значение
Значение пресмыкающихся для человека сравнительно невелико. Кожа крокодилов, крупных змей и ящериц применяется в кожевенной промышленности для изготовления чемоданов, ремней, обуви и т. п., однако эти предметы носят эксклюзивный характер, являясь предметом роскоши. Мясо и яйца многих черепах употребляются в пищу. Также могут употребляться в пищу некоторые ящерицы и змеи. Змеиный яд находит применение в медицине. Многие змеи полезны истреблением грызунов, а ящерицы — насекомых. Некоторые виды пресмыкающихся держат в качестве домашних животных. Аллигаторы, кайманы и некоторые крокодилы иногда разводятся на специальных фермах, для получения мяса, яиц и кожи.
Большую опасность для людей представляют ядовитые змеи, особенно в тропических странах. Многие крокодилы опасны для человека, наносят значительный ущерб животноводству. Пресноводные черепахи вредят рыбному промыслу. Крупные вараны, питоны и удавы так же могут быть опасны, будучи способными травмировать или даже убить человека.
Модельные объекты
В 2011 году был расшифрован геном пресмыкающегося — ящерицы анолис каролинский. Таким образом, это пресмыкающееся вошло в круг модельных объектов генетики.
См. также
Литература
- Наумов Н. П., Карташев Н. Н. Часть 2. Пресмыкающиеся, птицы, млекопитающие // Зоология позвоночных. — М.: Высшая школа, 1979. — С. 272.
- Карташев Н. Н., Соколов В. Е., Шилов И. А. Практикум по зоологии позвоночных. — М.: Высшая школа, 1981.
- Биологический энциклопедический словарь / под редакцией М. С. Гилярова и др. — М.: Советская Энциклопедия, 1989.
- Жизнь животных. Энциклопедия в шести томах. Том 5. (Земноводные. Пресмыкающиеся). Общая редакция члена-корреспондента АН СССР профессора Л. А. Зенкевича. — Москва: Просвещение, 1969. — 488 с.
- Phylonyms: A Companion to the PhyloCode (англ.) / de Queiroz K., Cantino P. D., Gauthier J. A., eds. — Boca Raton: Taylor & Francis Group, CRC Press, 2020. — 1352 p. — ISBN 978-1-138-33293-5.
Ссылки
- Пресмыкающиеся // Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров. — 3-е изд. — М. : Советская энциклопедия, 1969—1978.
- Классификация современных пресмыкающихся Архивная копия от 13 февраля 2012 на Wayback Machine (англ.)
| Таксономия | ||||
|---|---|---|---|---|
|
| ||||