
Таксо́н (лат. taxon, мн. ч. taxa; от др.-греч. τάξις — «порядок, устройство, организация») — группа в классификации, состоящая из дискретных объектов, объединяемых на основании общих свойств и признаков. Классификационные системы, использующие понятие таксона, обычно носят иерархический характер; применяются они в языкознании, библиографии и других науках, но прежде всего в биологии, а именно — в биологической систематике.
Учение о принципах и практике классификации и систематизации называется таксономией. Одна из наиболее фундаментальных проблем таксономии — это проблема способа бытия таксона, по-разному трактуемая в рамках различных исследовательских программ[значимость факта?].
Иерархические классификационные системы
При представлении знаний в виде некоторой классификационной системы изучаемые объекты группируются в классы (классификационные таксоны) с учётом ряда специально выбранных признаков (оснований классификации).
Делению на части (таксоны) класса классифицируемых объектов предшествует выделение основного класса объектов — таксономического универсума. Для биологии в целом это — совокупность живых организмов (особей), для сравнительно-исторического языкознания — совокупность идиолектов, для библиографии — совокупность печатных изданий; обычно же (при решении частных задач) роль таксономического универсума играет тот или иной таксон, уже выделенный в рамках более широкой классификационной системы. Говорят, что на этом универсуме задана таксономия, если выделена некоторая совокупность подклассов данного универсума (среди которых находится сам универсум (наибольший таксон), а пересечение таксонов всегда вновь образует некоторый таксон.
Выделению отдельных таксонов должно предшествовать и решение такого вопроса: какие группы могут считаться таксономически законными. Облик строящейся классификационной системы во многом предопределяется тем, как решается данный вопрос.
Классификационная система называется иерархической, если любые два входящих в неё таксона либо не пересекаются (нет объектов, одновременно принадлежащих обоим таксонам), либо находится в отношении соподчинения (то есть один из этих таксонов есть часть другого, причём не исключается возможность совпадения совокупностей объектов, образующих данные таксоны). Так, в систематике позвоночных классы Птицы (Aves) и Млекопитающие (Mammalia) — это непересекающиеся таксоны, а отряд Трубкозубые (Tubulidentata) есть часть объемлющего таксона Млекопитающие; при этом данный отряд являетсямонотипическим таксоном: включает один ныне живущий вид Orycteropus afer (трубкозуб африканский) (так что любая особь, входящая в таксон Tubulidentata, входит и в таксон O. afer, и наоборот). Таким образом, каждый таксон иерархической системы, с одной стороны, состоит из одного или большего числа таксонов более низкого уровня общности, с другой стороны — является в то же время частью другого таксона, относящегося к более высокому уровню общности (устроенная подобным образом система именуется также таксономической иерархией, а различные её уровни — таксономическими рангами).
Установление иерархии таксономических групп может проводиться по-разному. В биологии и лингвистике такая иерархия определяется степенью общности таксономических признаков, сформировавшихся на разных этапах эволюции таксономического универсума. Сами признаки при этом, вообще говоря, не находятся в отношениях подчинения, но возникают и развиваются на разных исторических этапах. Соответственно и каждый из выделяемых таксонов имеет своё определённое время существования.
Далее рассматриваются только иерархические классификационные системы, хотя встречаются и другие таксономические структуры — например, фасетные.
При классифицировании удобно выделить минимальные таксоны (виды) и перейти от таксономического универсума к рассмотрению классификационного поля — совокупности видов, которая, как правило, образует чёткое множество (в биологии роль минимальных таксонов обычно играют биологические виды, в сравнительно-историческом языкознании — языки). После такого перехода все остальные таксоны — объемлющие минимальные — можно трактовать уже не как совокупности классифицируемых объектов, а как множества минимальных таксонов.
Минимальные таксоны тем ближе друг к другу, чем «меньше» тот общий таксон, которому они одновременно принадлежат. Так, украинский язык ближе к русскому (они входят в одну и ту же восточнославянскую подгруппу славянской группы), чем к польскому (относящемуся к западнославянской подгруппе); поэтому между носителями русского и украинского языков возможно непосредственное общение без помощи переводчика, а с носителями польского взаимопонимание затруднено, хотя при попытке общения сходство языков живо ощущается.
Иерархическая классификационная система не всегда бывает доведена до уровня минимальных таксонов. При её построении нередко оперируют терминальными таксонами, которые могут соответствовать и таксонам надвидового ранга. Так, у А. Л. Тахтаджяна система покрытосеменных растений доведена до родов, которые и играют в ней роль терминальных таксонов (для некоторых наиболее обширных семейств даётся неполный перечень родов, что всегда явно оговаривается).
Таксоны в биологии
В Международном кодексе номенклатуры водорослей, грибов и растений (Шэньчжэньский кодекс, 2018) под термином «таксон» понимается таксономическая группа любого ранга. При этом подразумевается, что каждое растение рассматривается как принадлежащее к неопределённому числу таксонов последовательно соподчинённого ранга, среди которых ранг вида считается основным. Аналогично определяется таксон и в зоологии.
Представления о том, что такое «таксон», существенно различаются в двух основных направлениях современной филогенетической систематики. В рамках кладистики, вышедшей в 1970-е годы на первый план, господствует положение о том, что таксонами следует считать только монофилетические группы, иначе называемые кладами; приверженцы эволюционной таксономии признают законными также и группы парафилетические. Полифилетические группы следует удалять из классификационной системы в любом случае.
Характеристики биологического таксона
В качестве наиболее существенных характеристик (атрибутов) таксона в биологической систематике рассматриваютдиагноз, ранг и объём. По мере изменения классификации характеристики таксонов могут изменяться (в разных системах, например, таксоны одинакового объёма могут иметь разные диагнозы, или разные ранги, или же занимать в системе иное место).
При выделении таксонов часто рекомендуют, помимо принципа исключения полифилии, следовать принципу оптимальной диагностируемости (который учитывают при определении объёма и границ каждого таксона) и принципу единства уровня разнообразия и обособленности таксонов одинакового ранга. На практике эти два принципа соблюдают далеко не всегда.
Многие современные приверженцы кладистики вообще не определяют ранги для выделяемых ими групп и даже не приводят диагнозы, ограничиваясь построением кладограмм, дающих представление о филогении данных групп и их объёме.
Диагноз таксона
Под диагнозом таксона в биологической систематике понимают список его существенных признаков, то есть признаков, характеризующих таксон в такой степени, чтобы можно было отграничить его от других таксонов (которые могут содержать данный таксон, или же сами содержаться в нём, или, наконец, вообще с ним не пересекаться). Диагноз выступает как интенсиональное определение таксона, причём такое определение может быть монотетическим или политетическим.
Монотетическое определение таксона предусматривает, что каждый представитель таксона наделён определённым признаком (или совокупностью скоррелированных признаков), однозначно характеризующим данный таксон. Политетическое определение таксона предполагает, что такой общий признак указать нельзя, а таксон характеризуется некоторой группой признаков, причём ни одному из членов таксона нельзя приписать диагностирующие значения всех признаков, входящих в данную группу.
Автор, вводящий новый таксон и дающий ему название, должен следовать принципу действительного обнародования названия, то есть публиковать название, сопровождая его соответствующим диагнозом (даваемым по-латыни), только в распространённых периодических изданиях или книгах. Принцип предполагает, что автор названия должен позаботиться о том, чтобы все заинтересованные лица могли прочесть и понять его публикацию (без этого — по крайней мере в теории — любые названия никакой ценности для биологов не имеют и рассматриваться ими не будут).
Ранг таксона

Ранг в биологической систематике рассматривается как уровень в иерархии таксонов. Каждому такому уровню отвечает определённая таксономическая категория — совокупность таксонов данного ранга.
Представление о системе рангов как универсальных уровней иерархии, имеющих собственные названия, вошло в биологическую систематику в конце XVII века, когда в работах Августуса Бахмана (Ривинуса) (1690) и Жозефа Питтона де Турнефора (1694) была введена система соподчинённых таксономических категорий (наиболее детальной была система рангов Питтона де Турнефора: класс — секция — род — вид). При этом в работах данных учёных были чётко разграничены категории рода и вида и впервые применён принцип «один род — одно название». С тех пор ранги — несмотря на выдвигаемые рядом систематиков аргументы за их отмену — составляют неотъемлемую часть таксономической практики.
В классификации «отца систематики» Карла Линнея, опубликованной в его знаменитой книге Systema Naturae (1-е издание — 1735 г.; 10-е издание, условная дата опубликования которого принято за исходный пункт зоологической номенклатуры, — 1758 г.), имелось уже 6 рангов. Систему рангов Линнея проиллюстрируем на примере вида Homo sapiens (опуская последний уровень иерархии, точка зрения на который со времён Линнея изменилась кардинальным образом):
-
Царство (лат. regnum) — Animalia (животные)
-
Класс (лат. classis) — Mammalia (млекопитающие)
-
Порядок (Отряд) (лат. ordo) — Primates (приматы)
-
Род (лат. genus) — Homo (человек)
-
Вид (лат. species) — Homo sapiens (человек разумный)
- Разновидность (лат. varietas)
-
Вид (лат. species) — Homo sapiens (человек разумный)
-
Род (лат. genus) — Homo (человек)
-
Порядок (Отряд) (лат. ordo) — Primates (приматы)
-
Класс (лат. classis) — Mammalia (млекопитающие)
Заметим, что все рассмотренные в примере названия таксонов используются и поныне (валидны). Разумеется, в связи с прогрессом в развитии представлений учёных-систематиков о структуре биологического разнообразия число рангов со времён Линнея значительно возросло и к линнеевским категориям добавились тип, подтип, подкласс, семейство и др.
Для таксонов группы семейства форма, которую должно иметь название таксона, определяется рангом. Так, в зоологии для надсемейства, семейства, подсемейства и трибы название таксона должно оканчиваться соответственно на -oidea, -idae, -inae, -ini. В ботанике используется другой набор окончаний: -acea, -aceae, -oideae, -eae.
Относительное указание рангов содержательнее абсолютного. Первое (относительное) отражает объективные представления о соподчинённости групп. Например, утверждение о том, что к семейству животных A-idae относятся роды A-us, B-us и C-us, по сути говорит о том, что три группы объединены в четвёртую, и в принятой системе рангов речь идёт об объединении трёх родов в семейство. Второе (абсолютное) нередко отражает лишь субъективные представления о степени обособленности данной группы от других. Например, утверждение о том, что группу в составе A-us, B-us и C-us следует считать именно семейством A-idae, а не подсемейством A-inae, в отрыве от обсуждения вопроса о положении и иерархической соподчинённости этой группы в системе выглядит бессмысленным.
Объём таксона
Объём таксона может быть объективно задан путём перечисления входящих в него таксонов более низкого ранга. С течением времени представления систематиков об объёме конкретного таксона могут меняться — как по причине изменения взглядов на филогению рассматриваемой группы, так и в связи с обнаружением новых видов. В случае, когда широкое распространение получили одновременно две трактовки объёма таксона (узкая и широкая), во избежание путаницы пользуются латинскими выражениями sensu stricto (в «узком» смысле) и sensu lato (в широком смысле). Эти выражения добавляют к названию таксона, сокращая их обычно до s. str. и s. l. соответственно.
Например, выражение Arthropoda s. str. относится к группе, объединяющей только те таксоны, которые несомненно принадлежат к членистоногим (трилобитообразные, хелицеровые, ракообразные, многоножки, насекомые), в то время как выражение Arthropoda s. l. может относиться к группе, охватывающей, помимо перечисленных, таксоны не вполне ясного систематического положения: тихоходок и первичнотрахейных.
Заметим, что грамматическое оформление названий групп низшего ранга (вид, род, семейство) достаточно жёстко регулируется номенклатурными кодексами и потому в первую очередь связывается с рангом.
Что касается прочих названий, то их оформление правилами обычно не регулируются, и на практике происходит стихийное разделение этих названий на ранговые и объёмные: при изменениях классификации (когда меняется ранг или объём таксона) первые сохраняют за таксоном определённого ранга независимо от его объёма, а вторые — за таксоном определённого объёма независимо от его ранга. Например, в систематике членистоногих ранговым является название Insecta: оно всегда относится к таксону, имеющему ранг класса, а по объёму этот таксон менялся от Arthropoda (= Insecta sensu Linnaeus, 1758) до Pterygota (= Insecta sensu Latreille, 1810). С другой стороны, название Hexapoda Blainville, 1816 является объёмным и привязано к конкретной группе отрядов.
Отношения между биологическими таксонами
Как отмечалось при обсуждении иерархических классификационных систем, для любых двух таксонов системы могут встретиться две ситуации: 1) эти таксоны не пересекаются; 2) один из них является объемлющим, а второй — подчинённым.
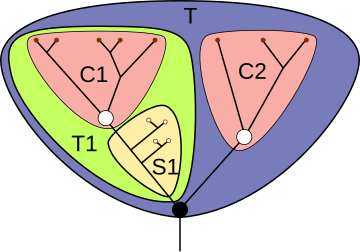
образующие объемлющую группу A
В кладистике важное значение имеет понятие сестринских групп (используемое и в эволюционной таксономии). «Сестринскими» называются две монофилетические группы, восходящие к общему предковому виду и образующие вместе с ним объемлющую монофилетическую группу. Если обе группы рассматриваются как таксоны, то говорят о сестринских таксонах; так, сестринскими таксонами являются отряды Rhynchocephalia (клювоголовые) и Squamata (чешуйчатые) класса пресмыкающихся.
Каждая из сестринских монофилетических групп естественным образом подразделяется на две подгруппы (сама же она выступает при этом как тотальная группа). Одну из указанных подгрупп, включающую всех потомков общего предка рецентных (ныне живущих) видов данных сестринских групп, Р. Джеффрис назвал в 1979 г. краун-группой (англ. crown group, в русскоязычной научной литературе встречается термин «кроновая группа»); вторая, названная В. Хеннигом стем-группой (англ. stem group, в русскоязычной научной литературе встречается термин «стволовая группа»), включает все вымершие формы данной группы за вычетом тех, которые вошли в краун-группу. Например, в отряде Rhynchocephalia краун-группа сводится к рецентному роду Sphenodon (гаттерия) и является таксоном; стем-группа парафилетична и включает вымершие роды семейства Sphenodontidae (клинозубые), а также представленные только ископаемыми формами семейства Gephyrosauridae и Pleurosauridae. На рисунке, приведённом справа, показана ситуация, когда у одной из сестринских групп стем-группа отсутствует (это значит, что ископаемые формы, которые могли бы в неё входить, ещё не обнаружены).
В эволюционной таксономии имеет место ещё один вид взаимоотношений между таксонами. Если некоторый парафилетический таксон образован вычитанием из монофилетической группы некоторых её монофилетических подгрупп, выделенных в отдельные таксоны, то он называется анцестральным (предковым) таксоном, а упомянутые подгруппы — производными (дочерними) таксонами; так, в рамках этого подхода класс Synapsida — анцестральный таксон для производного класса Mammalia.
Монофилия и парафилия в биологии
Приверженцы кладистики, как уже отмечалось, считают таксономически законными лишь таксоны, удовлетворяющие требованию монофилии; это означает, что таксону должны принадлежать его предок и все потомки данного предка. Предполагается, что только такой таксон является естественным, порождённым в процессе эволюции.
Для кладистики различные мнения относительно того, что собою представляют парафилетические и полифилетические группы, не имеют какого-либо значения. Сами эти группы не входят в круг её интересов.
Эволюционная таксономия признаёт законность рассмотрения в качестве таксонов и парафилетических групп (но в каждом конкретном случае не все такие группы могут одновременно обрести статус таксона). При этом парафилетическую группу можно определить как группу, получаемую из монофилетической группы путём вычитания из последней одной или нескольких монофилетических подгрупп. В такую группу входит её общий предок, но не все его потомки ей принадлежат.
Например, парафилетической является группа Рыбы (Pisces), получающийся из инфратипа Челюстноротые (Gnathostomata) вычитанием входящей в неё меньшей по объёму клады — надкласс Четвероногие (Tetrapoda).
Оба направления биологической систематики сходятся в том, что в качестве таксонов не должны рассматриваться полифилетические группы (в истории систематики такие группы возникали как продукт объединения по кажущейся схожести таких групп, сходство которых в действительности представляло собой результат конвергентной эволюции групп, в действительности не обладающих родством).
Надо заметить, что в некоторых группах организмов филогенетические отношения окончательно не установлены (а поэтому с достоверностью говорить об их моно-, пара- или полифилии рано). Построению адекватной филогении, помимо неполноты имеющихся данных, часто мешает наличие гомологических рядов, которое маскирует родственные отношения и обычно наиболее сильно выражено на уровнях родов, триб, подсемейств.
Для наглядного представления филогенетических взаимоотношений между различными таксонами в кладистике принято использовать кладограммы, отображающие иерархию монофилетических групп; при этом терминальным таксонам отвечают терминальные элементы кладограммы, нетерминальным — её рёбра (то есть междоузлия — вообще говоря, не все, если не всякая клада получает в данной системе статус таксона). Узловые точки кладограммы не соотнесены никаким таксонам: они соответствуют анцестральным (предковым) видам отдельных нетерминальных таксонов, причём такие виды всегда трактуются как гипотетические и не специализированы.
В эволюционной таксономии для наглядного представления таксономической системы используют — наряду с кладограммами — и филогенетические деревья. В них таксонам соотнесены и некоторые узловые точки (речь при этом идёт о парафилетических таксонах, играющих для других таксонов роль анцестральных групп). Филогенетическое дерево уже не находится в однозначном соответствии с филогенезом; требуется лишь, чтобы структура такого дерева не противоречила филогенезу.
Таксоны в лингвистике
В основе современной систематики языков лежитгенетическая классификация языков, строящаяся на основании определения родственных связей между ними, то есть на основе общего происхождения из предполагаемого праязыка. При этом языковое родство трактуется как общее свойство двух или нескольких языков, заключающееся в том, что их исконные минимальные значимые элементы (корневые морфемы и аффиксы) находятся в строго определённых соответствиях, отражающих регулярный характер звуковых преобразований материального фонда, восходящего к общему праязыку.
Лингвисты отмечают: «Генетическая классификация языков — аналог биологической классификации видов. Она позволяет систематизировать всё множество известных человеческих языков, которых насчитывается около 6000, сведя их к сравнительно небольшому числу языковых семей». Разработка генетической классификации языков — одна из важнейших задач сравнительно-исторического языкознания.
Таксономические категории в лингвистике
При генетическом подходе к языковой классификации оперируют таксономическими категориями: «семья», «ветвь», «группа» и т. п.; для группировок, включающих сильно расходящиеся языки, применяется также термин «макросемья». Впрочем, о полной стандартизации рангов лингвистических таксонов говорить рано: ничего подобного Международному кодексу ботанической номенклатуры в языкознании до сих пор не создано, и в терминологической практике объём лингвистического таксона изменчив: одно и то же объединение родственных языков может именоваться и «группой», и «семьёй». Тем не менее такие таксономические единицы, как макросемья — семья — группа — подгруппа — язык (а также наречие — диалект — говор), следует признать достаточно установившимися; в частности, их регулярно применяют этнологи, положившие несколько модифицированную классификацию языков в основу этнолингвистической классификации этносов.
Сходство родственных языков проявляется на всех уровнях структуры языка. На уровне базисной лексики (входящей в 100-словный список Сводеша) диалекты одного языка обычно имеют более 95 % совпадений. Языки, входящие в одну группу, имеют обычно 75—85 % совпадений. Языки, входящие в одну семью, обнаруживают 15—30 % совпадений (в промежуточном случае — 40—60 % совпадений — соответствующая общность языков может трактоваться и как семья, и как группа). Наконец, языки, входящие в одну макросемью, имеют примерно 5—10 % совпадений (в данном случае говорят о дальнем родстве, и для его обоснования обычно прибегают к ступенчатой реконструкции праязыка).
Попытка ввести в лингвистику более широкий набор таксономических категорий была предпринята С. Вурмом и К. Макэлханоном, предложившими в 1975 г. детальную классификацию папуасских языков. В качестве основных таксономических категорий в ней выступали: фила (языки, входившие в неё, имели 5—12 % совпадений в основном словарном фонде), ствол (12—28 % совпадений), семья (более 28 % совпадений); кроме того, встречались и промежуточные категории — субфилы, суперстволы. Особенностью классификации стало применение в ряде случаев двойных обозначений ранга у таксона. Например, среди стволов, составляющих филу Торричелли, мы находим семью-ствол Мариенберг и изолят-ствол урим. В первом случае степень родства между языками в составе группировки — такая же, как между языками одной семьи, но в состав рассматриваемой филы данная группировка входит на правах самостоятельного ствола; во втором случае ствол состоит из одного изолированного (в пределах филы Торричелли) языка урим (или калп). Впрочем, дальнейшего развития в рамках генетической классификации языков двойные обозначения ранга не получили.
Языковые группы
Представление о языковых группах возникает в XVII в. и восходит к небольшой работе «Краткое рассуждение о языке европейцев», написанной в 1599 г. в Голландии французским учёным-филологом И. Ю. Скалигером и опубликованной во Франции в 1610 году уже после смерти учёного. В данной работе Скалигер установил для европейских языков 11 «языков-матерей» (linguae matrices, то есть праязыков), к которым восходят новые европейские языки (в современной терминологии, речь идёт о таких группах: 1) греческая; 2) романская; 3) германская; 4) славянская; 5) албанская; 6) тюркская; 7) угорская; 8) финно-пермская; 9) гойдельская; 10) бриттская; 11) баскская). Сами же эти matrices, по Скалигеру, «не связаны между собой никакими узами родства».
Языковые семьи
Вопрос о более отдалённом родстве между языками (чем родство между языками, входящими в одну группу) был поставлен в XVIII в. Так, голландский учёный Л. тен Кате в работе «Введение в изучение благородной части нижненемецкого языка» (1723) уже предполагал родство между 7 из 11 matrices Скалигера (1—5 и 9—10), предвосхищая тем самым понятие об индоевропейской семье (но он имел дело лишь с языками Европы).
Российский учёный-энциклопедист М. В. Ломоносов в материалах и черновых набросках к «Российской грамматике» (вышла в 1755 г.) последовательно противопоставлял родственные языки неродственным, а близкородственные — дальнеродственным (и ввёл тем самым представление об иерархии в классификации языков, находящей ныне своё выражение в иерархии лингвистических таксонов). Он сформулировал положение об образовании родственных языков путём последовательного дробления языка, относящегося к более ранней эпохе, и показал, что русский язык обладает наибольшим родством с другими славянскими языками, более отдалённым — с «курляндским» (латышским, то есть с балтийскими), ещё более отдалённым — с латинским, греческим и немецким. Ломоносов писал: «Представим долготу времени, которого сии языки разделились! Подумай же, когда курляндский! Подумай же, когда латинский, греческий, немецкий, российский! О глубокая древность». В то же время учёный отрицал какое-либо родство упомянутых языков с финским, «мексиканским» (ацтекским), готтентотским, китайским.
Очередной шаг сделал английский востоковед языковед У. Джонс, который в 1786 году декларировал, выступая в Калькутте с публичным докладом, родство санскрита и древнеперсидского с важнейшими древними и современными языками Европы. Наконец, немецкий лингвист Ф. Бопп стал в 1816 году создателем сравнительного языкознания, научно обосновав существование индоевропейской языковой семьи, в которую включил санскрит, персидский и некоторые (позднее — большинство) языков Европы. Для данного объединения языков немецкий востоковед и полиглот Ю. Клапрот в 1823 г. предложил название «индогерманские языки», позднее вытесненное современным названием «индоевропейские языки».
За сорок лет до этого возникло представление о существовании другой хорошо известной языковой семьи — семитской семьи: уже в 1781 году немецкий историк и филолог А. Л. фон Шлёцер объединил ряд мёртвых и живых языков Ближнего Востока в общность семитских языков (правда, данная общность длительное время трактовалась как группа). В 1836 году немецкий лингвист В. Гумбольдт обосновал существование малайско-полинезийской семьи, которую австрийский лингвист и этнограф В. Шмидт в 1899 г. предложил переименовать в австронезийскую семью (ныне используются оба названия, причём малайско-полинезийская семья трактуется как крупнейшее подразделение более широкого объединения — австронезийской надсемьи). Ещё одна крупная языковая семья — китайско-тибетская; её выделил (под названием «индо-китайской») немецкий китаевед и лингвист А. Конради в 1896 году (причём ошибочно включил в её состав тайские языки — ошибка, исправленная американским антропологом и лингвистом П. Бенедиктом в 1942 г.), а современное название семьи впервые употребил Ж. Пшилюский в 1924 г.
Языковые макросемьи
Таксономической категорией наивысшего ранга в лингвистике является категория макросемьи. Среди макросемей старейшей по времени выделения является афразийская макросемья (первоначально ей придавали, впрочем, ранг семьи). Представление о существовании данной языковой общности, в которую наряду с семитскими языками входит также и ряд языков Северной Африки, складывается в середине XIX в., восходя к немецкому египтологу К. Лепсиусу (который в 1863 году объединил в одну семью семитские, кушитские, берберские языки, а также древнеегипетский и язык хауса) и австрийскому лингвисту Ф. Мюллеру, предложившему в 1876 г. термин «семито-хамитские языки»; утвердилось данное представление под влиянием работ австрийского египтолога Л. Райниша.
В 1955 г. американский лингвист Дж. Гринберг предложил заменить термин «семито-хамитские языки» на «афро-азиатские языки» (англ. Afro-Asiatic languages); в отечественном языкознании ныне принят предложенный И. М. Дьяконовым в 1974 г. термин «афразийские языки». С конца XX в. афразийские языки рассматриваются как макросемья.

Однако последовательное изучение дальнего языкового родства относится уже к XX веку. В 1910-х гг. датский лингвист Х. Педерсен выдвинул идею о существовании ностратической макросемьи; детальная разработка ностратической теории началась позже — в 1960-е гг. (работы советских компаративистов В. М. Иллич-Свитыча и А. Б. Долгопольского). В 1980-х гг. советский компаративист С. А. Старостин обосновал существование сино-кавказской макросемьи.
В приложении 1 к книге С. А. Бурлак и С. А. Старостина «Сравнительно-историческое языкознание» все языки мира объединены в 11 макросемей (плюс несколько языков-изолятов); оговаривается, что некоторые из этих макросемей пока представляют собой сугубо гипотетические объединения. Лингвисты считают, что возраст макросемей может лежать в пределах от 10 000 до 30 000 лет; вопрос о том, восходят ли все они к единому общему прамировому языку («гипотеза моногенеза») или же возникли независимо, остаётся открытым.
В начале XXI века появляется всё большее число исследований, в которых для установления филогенетических связей между лингвистическими таксонами применяются те же алгоритмы вычислительной филогенетики, что и при построении современных биологических классификаций. В качестве примера приведём некоторые результаты выполненного в 2015 г. анализа совокупности около 1000 языков и диалектов Евразии, принадлежащих к различным семьям и макросемьям. В качестве анализировавшейся совокупности признаков было использовано подмножество из 40 наиболее устойчивых лексем, взятых из списка Сводеша (в фонетической записи). Анализ подтвердил существование большинства выделяемых традиционными методами группировок, хотя выявились и неожиданные результаты. Среди них: чукотско-камчатская семья оказалась сестринской группой для индоевропейской семьи; японский язык группировался с айнским и аустроазиатскими, а не с другими алтайскими; дравидийская семья не обнаружила никакого родства с другими ностратическими языками Впрочем, результаты данного и подобных ему исследований пока следует считать предварительными.
Монофилия и парафилия в лингвистике
При генеалогической классификации языков возникает та же проблема различения монофилетических и парафилетических групп, что и в биологической систематике. Целесообразность использования парафилетических групп в лингвистической систематике подкреплена тем обстоятельством, что процессы языковой дивергенции в ряде семей и групп достаточно полно отражены в письменных памятниках, так что нередко праязык оказывается не чисто гипотетическим, а вполне реальным языковым образованием. Так, В. Н. Топоров, описывая состав индоевропейской семьи, выделял 17 групп (часть из которых — монотипические таксоны, представленные одним языком). Среди этих таксонов одинаковый ранг группы имеют италийские и романские языки; между тем известно, что для романских языков праязыком служит один из италийских языков — латинский (более точно: протороманский совпадает с так называемой вульгарной латынью — сестрой классической латыни, общий предок которых — латынь Гракхов — относится ко второй половине II в. до н. э.).
Для наглядного представления взаимоотношения между монофилетическими лингвистическими таксонами в языкознании (как и в биологии) используют кладограммы (именуемые в лингвистике, впрочем, родословными или генеалогическими деревьями). Такой способ отображения филогении языковых таксонов восходит к 1861—1862 гг., когда немецкий учёный А. Шлейхер в своём «Компендиуме сравнительной грамматики индогерманских языков» представил в виде родословного древа развитие индоевропейских языков (он же осуществил первую реконструкцию индоевропейского праязыка). Приведём это древо (заменяя в ряде случаев шлейхеровские названия лингвистических таксонов на современные):
|
|||||||||||||||||||||||||||||||||||||||||||||||||
Заметим, что в современных классификациях индоевропейских языков (в которых добавились новые группы: анатолийская, тохарская, армянская, нуристанская и др.) сохранилась часть выделенных Шлейхером промежуточных таксонов, а вот представление об арио-греко-итало-кельтской и греко-итало-кельтской общностях было отброшено.
Генеалогические деревья весьма часто приводятся в современных изданиях, рассматривающих историю различных языковых семей.
Таксоны в библиографии
Важным примером классификации иерархического типа служит Универсальная десятичная классификация (УДК), используемая для индексирования книг и документов. Основные правила данной системы были разработаны американским библиографом М. Дьюи, который в 1876 г. предложил систематизировать библиотечные фонды на основе десятичной классификации идей и понятий (принцип такой классификации содержался ещё в проекте априорного языка, представленном в 1794 г. французским адвокатом и филологом Ж. Делормелем на рассмотрение Национального конвента Франции) и создал систему Десятичная классификация Дьюи. Сама система УДК, опубликованная впервые в 1897 г., возникла в результате переработки системы Дьюи бельгийскими библиографами П. Отле и А. Лафонтеном, создавшими в 1895 г. в Брюсселе Международный библиографический институт.
В систему УДК совокупность человеческих понятий и представлений разделена на 10 классов, обозначенных цифрами от 0 до 9 и играющим роль таксонов высшего ранга. Вся совокупность математических и естественных наук отнесена к классу 5; подобласти данной совокупности отвечают разделам этого класса, получившим индексы от 50 до 59, а ещё более узким подобластям отвечают более длинные десятичные индексы. Наиболее длинные индексы соответствуют минимальным таксонам — самым узким тематическим областям, различаемым в данной системе.
Выберем, например, в качестве минимального таксона уравнения Лагранжа (одно из основных понятий аналитической механики) и рассмотрим, какое место он занимает в системе УДК. Получаем следующую иерархическую лестницу таксонов:
- 5 — Математика и естественные науки
- 53 — Физика
- 531 — Общая механика. Механика твёрдого тела
- 531.3 — Динамика. Кинетика
- 531.31 — Основы динамики и кинетики
- 531.314 — Принцип Гамильтона. Уравнения Гамильтона. Принцип наименьшего действия
- 531.314.2 — Уравнения Лагранжа
По аналогичному принципу построены и другие современные системы библиографической классификации — в частности, используемая в России система Библиотечно-библиографическая классификация (ББК).
См. также
- Комментарии
- Источники
Литература
Общие вопросы таксономии
- Зуев В. В., Розова С. С. Проблема способа бытия таксона в биологической таксономии // Вопросы философии. — 2003. — № 2. — С. 90—103. Архивировано 17 апреля 2013 года.
- Мейен С. В., Шрейдер Ю. А. Методологические аспекты теории классификации // Вопросы философии. — 1976. — № 12. — С. 67—79.
- Шрейдер Ю. А., Шаров А. А. Системы и модели. — М.: Радио и связь, 1982. — 152 с. — (Кибернетика).
Биологическая таксономия
- Linnaeus, Carolus. Systema naturae per regna tria naturae: secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. — Holmia: Laurentius Salvius, 1758.
- Джеффри Ч. Биологическая номенклатура = Charles Jeffrey. Biological Nomenclature. Second edition / Пер. с англ. — М.: Мир, 1980. — 120 с. — 15 000 экз. — УДК 574:001.4
- Клюге Н. Ю. Система альтернативных номенклатур надвидовых таксонов // Энтомологическое обозрение. — 1999. — № 78 (1). — С. 224—243.
- Майр Э. Принципы зоологической систематики: Пер. с англ. = Ernst Mayr. Principles of Systematic Zoology. — М.: Мир, 1971. — 455 с. — УДК 590
- Павлинов И. Я., Любарский Г. Ю. Биологическая систематика: Эволюция идей. — М.: Т-во научных изданий КМК, 2011. — 667 с. — ISBN 978-5-87317-685-4.
- Шаталкин А. И. Биологическая систематика. — М.: Изд-во Моск. ун-та, 1988. — 184 с. — ISBN 5-211-00145-1.
- Шаталкин А. И. Таксономия. Основания, принципы и правила. — М.: Товарищество научных изданий КМК, 2012. — 600 с. — 600 экз. — ISBN 978-5-87317-847-6. — УДК 579.8.06
- Шипунов А. Б. Основы теории систематики. — М.: Книжный дом «Университет», 1999. — 56 с.
Лингвистическая таксономия
- Амирова Т. А., Ольховиков Б. А., Рождественский Ю. В. История языкознания. 2-е изд. — М.: Издат. центр «Академия», 2005. — 672 с. — ISBN 5-7695-2377-8.
- Бурлак С. А., Старостин С. А. Сравнительно-историческое языкознание. — М.: Издат. центр «Академия», 2005. — 432 с. — ISBN 5-7695-1445-0.
- Сравнительно-историческое изучение языков разных семей. Задачи и перспективы. — М.: Наука, 1982. — 343 с.
- Широков О. С. Введение в языкознание. — М.: Изд-во Моск. ун-та, 1985. — 264 с.
Библиографическая таксономия
- Дрезен Э. К. За всеобщим языком. Три века исканий. 3-е изд. — М.: Едиториал УРСС, 2012. — 272 с. — ISBN 978-5-354-01430-9.
Ссылки
- Универсальная десятичная классификация (УДК). Таблицы официального российского эталона Универсальной десятичной классификации (УДК) по состоянию на 2001 год
| В библиографических каталогах |
|---|