

Протеасо́ма (от англ. protease — протеиназа и лат. soma — тело) — многобелковый комплекс, разрушающий ненужные или дефектные белки при помощи протеолиза (химической реакции, при которой происходит разрыв пептидных связей) до коротких пептидов (4—25 аминокислотных остатков). Эти пептиды затем могут быть расщеплены до отдельных аминокислот. Протеасомы присутствуют в клетках эукариот, архей и некоторых бактерий. В эукариотических клетках протеасомы содержатся и в ядре, и в цитоплазме. Деградация 80—90 % внутриклеточных белков происходит при участии протеасомы. Для того чтобы белок-мишень расщепился протеасомой, он должен быть помечен путём присоединения к нему маленького белка убиквитина. Реакция присоединения убиквитина катализируется ферментами убиквитинлигазами. Присоединение первой молекулы убиквитина к белку служит для убиквитинлигаз сигналом для дальнейшего присоединения молекул убиквитина. В результате к белку оказывается присоединена полиубиквитиновая цепь, которая связывается с протеасомой и обеспечивает расщепление белка-мишени. В целом вся эта система получила название убиквитин-зависимой деградации белка.
Протеасомальная деградация белка важна для протекания многих клеточных процессов, включая клеточный цикл, регуляцию экспрессии генов и ответ на окислительный стресс. В 2004 году Аарон Чехановер, Аврам Гершко и Ирвин Роуз были удостоены Нобелевской премии по химии «за открытие убиквитин-зависимой деградации белка».

История открытия
До открытия убиквитин-зависимой системы деградации белков считалось, что деградация белков в клетке происходит, главным образом, за счёт лизосом. Лизосомы — это мембранные органеллы с кислой внутренней средой, содержащей протеазы. Они способны утилизировать экзогенные белки, захваченные клеткой в процессе эндоцитоза, белки, связанные с мембранами, и повреждённые органеллы. Однако в 1977 году Альфред Голдберг доказал существование АТФ-зависимой системы деградации белка в ретикулоцитах, которые лишены лизосом. Это позволило предположить, что существует как минимум ещё один механизм внутриклеточного расщепления белков. В 1978 году было показано, что соответствующая протеаза состоит из полипептидных цепей нескольких типов. Кроме того, при исследовании посттрансляционных модификаций гистонов была обнаружена неожиданная ковалентная модификация: присоединение к боковой цепи лизина в гистоне C-концевого остатка глицина убиквитина — небольшого белка с неизвестной функцией. Было установлено, что описанный ранее АТФ-зависимый фактор протеолиза 1 и убиквитин являются одним и тем же белком. В дальнейшем АТФ-зависимый белковый комплекс, ответственный за убиквитин-опосредованную деградацию белка, был выделен из лизата клеток и назван 26S-протеасомой.
Большая часть ранних работ, которые впоследствии привели к открытию протеасомной системы деградации белков, была выполнена в конце 1970-х — начале 1980-х годов в лаборатории Аврама Хершко в Технионе, где Аарон Чехановер был аспирантом. Ключевые концептуальные идеи Хершко выработал за год работы в лаборатории Ирвина Роуза, хотя Роуз впоследствии и преуменьшал свою роль в открытии. Все трое разделили Нобелевскую премию по химии в 2004 году за открытие этой системы.
Хотя электронно-микроскопические данные, указывающие на то, что структура протеасомы представляет собой несколько колец, уложенных в стопку, были доступны уже в середине 1980-х годов, первая структура коровой части протеасомы, составленная на основе данных рентгеноструктурного анализа, была получена только в 1994 году.
Структура


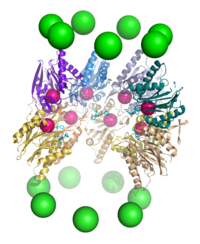
Компоненты протеасомы часто называются в соответствии с их коэффициентами седиментации в сведбергах (обозначается буквой S). Протеасома, активная в расщеплении белков, называется 26S-протеасомой и обычно состоит из коровой 20S-протеасомы и одной или двух регуляторных частиц 19S (PA700) и 11S, которые присоединяются к торцам коровой частицы. Хотя присоединение двух регуляторных частиц, строго говоря, приводит к формированию протеасомы с коэффициентом седиментации 30S, термин «30S-протеасома» в литературе практически не используется, а название «26S-протеасома» применяется по отношению к обеим изоформам. Кроме регуляторной частицы 19S, в состав 26S-протеасомы могут входить и другие регуляторные компоненты: PA28α/β (11S REG), PA28γ (REGγ), PA200, PI31. В состав некоторых протеасом входит другая регуляторная частица — 11S. Она взаимодействует с 20S-частицей так же, как 19S, и может принимать участие в деградации чужеродных белков, например, синтезированных во время вирусной инфекции.
Размеры протеасом относительно эволюционно стабильны и составляют 150 на 115 ангстрем. Внутренняя полость имеет максимальную ширину 53 ангстрема, однако вход в протеасому может иметь ширину всего 13 ангстрем, это указывает на то, что для входа в протеасому белок должен быть специфичноденатурирован.
20S-частица
20S-частицы протеасом прокариот и эукариот имеют принципиально одинаковую четвертичную структуру и состоят из 28 субъединиц, организованных в четыре семичленных кольца, уложенных друг на друга в виде стопки. Однако разнообразие субчастиц протеасомы зависит от конкретного организма: разнообразие субъединиц выше у многоклеточных организмов по сравнению с одноклеточными и у эукариот по сравнению с прокариотами. Протеасомы прокариот состоят из 14 копий идентичных α-субъединиц, которые формируют внешние кольца, и 14 копий идентичных β-субъединиц, которые формируют внутренние кольца. В эукариотической протеасоме все семь субъединиц одного кольца отличаются по структуре, то есть протеасома состоит из двух копий семи разных α-субъединиц и двух копий семи разных β-субъединиц. Несмотря на небольшие различия, с точки зрения пространственной структуры α- и β-субъединицы, тем не менее, очень похожи. α-субъединицы отвечают за присоединение к 20S-протеасоме регуляторных частиц, а их N-концевые участки прикрывают вход в полость протеасомы, что исключает неконтролируемый протеолиз. β-субъединицы имеют протеазные центры и являются каталитическими компонентами протеасомы. У архей, например, у Thermoplasma acidophilum, все β-субъединицы одинаковы, поэтому протеасома содержит 14 идентичных протеазных центров. В протеасомах млекопитающих каталитически активными являются только β1-, β2- и β5-субъединицы, причём все эти субъединицы обладают разными субстратными специфичностями (пептидил-глутамил-гидролизующей, трипсиноподобной и химотрипсиноподобной соответственно). В гематопоэтических клетках под действием провоспалительных сигналов, таких как цитокин интерферон γ, могут экспрессироваться альтернативные формы β-субъединиц, которые обозначаются β1i, β2i и β5i. Протеасому, содержащую эти альтернативные β-субъединицы, называют иммунопротеасомой, и её субстратная специфичность несколько отличается от таковой у обычной протеасомы. В середине 2010-х годов в клетках человека были идентифицированы необычные протеасомы, лишённые коровой субъединицы α3. В таких протеасомах (известных также как α4-α4 протеасомы) 20S-коровая часть содержит субъединицу α4 вместо α3. Альтернативные α4-α4 протеасомы были также выявлены у дрожжей. Хотя функции этой изоформы протеасомы неизвестны, клетки, экспрессирующие их, отличаются повышенной устойчивостью к токсичному действию ионов металлов, например, кадмия.
19S-регуляторная частица
У эукариот 19S-частица состоит из 19 отдельных белковых молекул, которые образуют 9-субъединичное основание, непосредственно взаимодействующее с α-кольцом 20S-коровой частицы, и 10-субъединичную «крышечку». Шесть из девяти белков основания являются АТФазами из семейства ААА, их гомологи имеются у архей и называются PAN (от англ. Proteasome-Activating Nucleotidase — нуклеотидаза, активирующая протеасому). Для взаимодействия 19S- и 20S-частиц необходимо, чтобы субъединицы 19S-частицы с АТФазной активностью были связаны с АТФ, и для протеасомной деградации уложенных и убиквитинированных белков необходим гидролиз АТФ. Строго говоря, гидролиз АТФ нужен только для денатурации белков, но само связывание с АТФ может способствовать протеканию других этапов разрушения белков (например, сборке комплекса, открыванию ворот, транслокации и протеолизу). Кроме того, связывание АТФ с АТФазами само по себе способствует быстрой деградации неуложенных белков. Хотя абсолютная необходимость в АТФ показана лишь для разрушения пространственной структуры белка, до конца не исключена возможность того, что гидролиз АТФ необходим для сопряжения разных этапов деградации белка.
В 2012 году две исследовательские группы независимо представили молекулярную структуру 26S-протеасомы, полученную с помощью электронной микроскопии единичных частиц. Позднее с помощью криоэлектронной микроскопии была построена атомная модель протеасомы. В центре 19S-частицы, недалеко от 20S-частицы, находятся ААА-АТФазы, которые образуют гетерогексамерное кольцо (Rpt1/Rpt2/Rpt6/Rpt3/Rpt4/Rpt5). Это кольцо представляет собой тример димеров Rpt1/Rpt2, Rpt6/Rpt3 и Rpt4/Rpt5. АТФазы димеризуются при помощи своих N-концевых биспиральных (англ. coiled coil) участков, которые выступают из гексамерного кольца. Два регуляторных белка Rpn1 и Rpn2, лишённые АТФазной активности, связываются с концами димеров Rpt1/2 и Rpt6/3 соответственно. Рецептор убиквитина Rpn13 связывается с Rpn2. Крышечка закрывает половину ААА-АТФазного гексамера (Rpt6/Rpt3/Rpt4) и взаимодействует непосредственно с 20S-частицей при помощи Rpn6 и, в меньшей степени, Rpn5. Субъединицы Rpn9, Rpn5, Rpn6, Rpn7, Rpn3 и Rpn12, структурно родственные друг другу, а также субъединицам комплексов COP9 и eIF3, объединяются в подковообразную структуру, в которой заключён гетеродимер Rpn8/Rpn11. Субъединица Rpn11, которая является деубиквитинирующим ферментом, помещается недалеко от внутренней полости гексамерного кольца из ААА-АТФаз, что идеально подходит для удаления убиквитиновых остатков непосредственно перед транслокацией разрушаемых белков в 20S-частицу. Второй из известных на данный момент убиквитиновых рецепторов, Rpn10, находится на периферии крышечки, рядом с субъединицами Rpn8 и Rpn9.
Конформационные изменения в 19S-частице
Для регуляторной частицы 19S известны три чётко различающиеся конформации. Вероятно, все они играют важную роль в распознавании и разрушении субстрата. Первая конформация характеризуется наименьшей энергией, что достигается расположением ААА-доменов АТФаз в виде лестницы или пружины. В присутствии АТФ, но в отсутствие субстрата, наблюдается вторая, менее распространённая конформация, которая отличается расположением крышечки относительно ААА-АТФазного модуля. В присутствии АТФ-γS или субстрата реализуется третья конформация с сильно изменённой структурой ААА-АТФазного модуля.
Регуляция 20S-частицы 19S-частицей
19S — регуляторная частица, она стимулирует разрушение субстрата 20S-субъединицей. Главной функцией 19S-частицы является открывание ворот в 20S, которые препятствуют входу субстратов внутрь протеасомы. Удалось определить механизм, по которому АТФазы открывают ворота 20S-частицы. Для открывания ворот необходим специфический мотив на C-концах АТФаз. За счёт него C-концы АТФаз входят в специальные карманы в верхней части 20S-частицы, закрепляя АТФазный комплекс на 20S-протеолитическом комплексе, благодаря чему часть протеасомы, ответственная за денатурацию субстрата, оказывается сопряжена с деградирующим модулем. Само связывание C-концов АТФаз с 20S вызывает открытие ворот в последней подобно тому, как ключ открывает замок. Структурные изменения, сопровождающие открытие ворот, также были изучены.
11S-регуляторная частица
С 20S-протеасомой может взаимодействовать ещё одна регуляторная частица, имеющая массу 11S и представляющая собой гептамер (она также известна как PA28 или REG). Она не содержит АТФаз и способствует разрушению коротких пептидов, но не больших белков. Вероятно, это связано с тем, что 11S-частица не может денатурировать большие белковые молекулы. Механизм взаимодействия 11S-частицы с 20S-протеасомой напоминает взаимодействие 19S-частицы с последней: 11S-частица связывается с 20S своим C-концевым хвостом и вызывает конформационные изменения в α-кольце, которые вызывают открытие ворот 20S-частицы. Экспрессия 11S-частицы запускается интерфероном γ, и эта частица, наряду с β-субъединицами иммунопротеасомы, ответственна за образование пептидов, которые связываются с главным комплексом гистосовместимости.
Сборка
Сборка протеасомы — очень сложный процесс, при котором множество отдельных белковых молекул должны объединиться с образованием активного комплекса. β-субъединицы синтезируются с N-концевыми «пропептидами», которые при сборке 20S-частицы подвергаются посттрансляционным модификациям, чтобы потом войти в состав каталитического активного центра. 20S-частица собирается из двух половин, каждая из которых содержит β-кольцо, состоящее из семи субъединиц и связанное с семичленным α-кольцом. Полная 20S-частица формируется при соединении двух половин посредством β-колец, которое сопровождается треонинзависимым автолизом пропептидов, в результате чего образуется активный центр протеасомы. Взаимодействие β-колец опосредовано соляными мостиками и гидрофобными взаимодействиями консервативных α-спиралей, причём мутации в них делают невозможной сборку протеасомы. Сборка каждой половины протеасомы начинается с образования гептамерного кольца из α-субъединиц, которое служит в качестве матрицы для сборки β-кольца. Механизм сборки α-кольца не изучен.
Регуляторная частица 19S собирается из двух частей — основания и крышечки. Сборка основания происходит при участии четырёх шаперонов Hsm3/S5b, Nas2/p27, Rpn14/PAAF1 и Nas6/ганкирин (первое название — у дрожжей, второе — у млекопитающих). Шапероны взаимодействуют с ААА-АТФазными субъединицами и обеспечивают формирование из них правильного гексамерного кольца. Сборке основания также способствует деубиквитинирующий фермент Ubp6/Usp14, однако он не является строго необходимым. Пока что неизвестно, связана ли сборка 19S-частицы со сборкой 20S-частицы. Крышечка собирается отдельно от основания без участия шаперонов.
Разрушение белков
Убиквитинирование

Белки, которые необходимо разрушить с помощью протеасом, метятся ковалентным присоединением маленького белка убиквитина к остаткам лизина. Присоединение убиквитина осуществляется тремя ферментами. На первом этапе убиквитин-активирующий фермент, известный как Е1, гидролизует АТФ и аденилилирует молекулу убиквитина. Далее аденилилированный убиквитин переносится на остаток цистеина фермента E1 одновременно с аденилированием второго убиквитина. Аденилилированный убиквитин далее переносится на остаток цистеина второго фермента — убиквитин-конъюгирующего фермента (Е2). На последнем этапе фермент из обширной группы убиквитинлигаз (Е3) распознаёт белок, который нужно разрушить, и переносит на него убиквитин с Е2. Таким образом, субстратную специфичность убиквитин-протеасомной системы обеспечивает именно Е3. Для распознавания крышечкой протеасомы белок должен нести цепочку из как минимум четырёх мономеров убиквитина (то есть быть полиубиквитинированным).
Механизм распознавания полиубиквитинированного белка протеасомой до конца не понятен. Рецепторы убиквитина имеют N-концевой убиквитин-подобный домен (англ. ubiquitin-like domain, UBL), а также один или несколько убиквитин-ассоциированных доменов (англ. ubiquitin-associated domain, UBA). UBL-домены распознаются крышечкой протеасомы, а UBA взаимодействует с убиквитином при помощи трёх α-спиралей. Белки-рецепторы убиквитина могут доставлять полиубиквитинированные белки к протеасоме, однако детали процесса и его регуляция неясны.
Сам убиквитин состоит из 76 аминокислотных остатков и получил своё название за повсеместное распространение (от англ. ubiquitous — «повсеместный»). Этот белок очень консервативен и выявляется у всех эукариот. Гены эукариот, кодирующие убиквитин, образуют тандемные повторы, вероятно, из-за того, что они очень активно транскрибируются, чтобы поддерживать необходимый уровень убиквитина в клетке. Высказываются предположения, что убиквитин — самый медленно эволюционирующий белок из известных. Убиквитин содержит семь остатков лизина, к которым могут присоединяться другие молекулы убиквитина, благодаря чему возможно образование полиубиквитиновых цепочек разного вида. Протеасома распознаёт полиубиквитиновые цепочки, в которых каждая следующая молекула убиквитина присоединяется к 48-му остатку предыдущего убиквитина, а все остальные участвуют в других клеточных процессах, то есть представляют собой посттрансляционные модификации.
Денатурация и транслокация
Полиубиквитинированный белок распознаётся 19S-субъединицей, и для его денатурации (то есть разрушения пространственной структуры) необходима энергия АТФ. Далее белок должен попасть внутрь 20S-субъединицы, а именно в её активный центр. Поскольку полость 20S-субъединицы очень узкая и закрыта воротами из N-концов субъединиц α-кольца, субстрат должен быть хотя бы отчасти денатурирован. Кроме того, с него должна быть снята убиквитиновая метка. Переход денатурированного белка внутрь активного центра протеасомы называется транслокацией. Однако порядок, в котором происходит денатурация и деубиквитинирование белков-субстратов, неизвестен. То, какой из этих этапов является скоростьлимитирующим, зависит от субстрата. Степень денатурации, которая позволяет субстрату попасть в активный центр, также неизвестна, однако третичная структура и некоторые связи внутри белковой молекулы, например, дисульфидные связи, препятствуют разрушению белка. Наличие неуложенных участков определённой длины внутри белка или на его конце облегчает эффективное разрушение.
Ворота, сформированные α-субъединицами, препятствуют попаданию внутрь 20S-частицы пептидов, состоящих из более чем четырёх остатков. Перед транслокацией пептида происходит его денатурация, для которой необходима энергия гидролиза АТФ, однако сам процесс транслокации не нуждается в дополнительном источнике энергии. Протеасома может разрушать денатурированные белки даже в присутствии негидролизуемого аналога АТФ, но не белки в нативной форме, что свидетельствует о необходимости энергии АТФ только для процесса денатурации. Проход денатурированного субстрата сквозь ворота идёт по типу облегчённой диффузии, если при этом АТФазные субъединицы 19S-частицы связаны с АТФ.
Денатурация глобулярных белков в целом протекает по одному и тому же механизму, хотя некоторые его реакции зависят от аминокислотного состава субстрата. Длинные участки, состоящие из повторяющихся остатков глицина и аланина, подавляют денатурацию, что снижает эффективность протеасомного разрушения, вероятно, из-за того, что гидролиз АТФ и денатурация оказываются разобщены. Результатом такого неполного разрушения являются частично разрушенные белки. Повторы глицина и аланина встречаются в природных белках, например, в фиброине шёлка. Кроме того, они имеются у некоторых белков вируса Эпштейна — Барр и нарушают работу протеасом, благодаря чему нарушается презентация антигенов на главном комплексе гистосовместимости и, следовательно, облегчается размножение вируса.
Протеолиз
Протеолиз субстрата β-субъединицами 20S-частицы происходит по типу треонинзависимой нуклеофильной атаки. Для депротонирования активной гидроксильной группы треонина может быть необходима вода. Деградация происходит в центральной полости протеасомы, которая образуется при взаимодействии двух β-колец и в норме не выпускает наружу частично разрушенные белки, разрушая субстрат до пептидов из 7—9 остатков (хотя их длина может варьировать от 4 до 25 остатков в зависимости от организма и субстрата). Чем определяется длина пептидов, образующихся при протеолизе в протеасоме, неизвестно. Хотя три β-субъединицы используют один и тот же механизм для деградации белков, они обладают слегка различающейся субстратной специфичностью, из-за чего их подразделяют на трипсин-подобные, химотрипсин-подобные и пептидил-глутамил-подобные. Специфичность обусловлена взаимодействиями атомов соседних аминокислотных остатков вблизи активного центра. Кроме того, каждая каталитическая β-субъединица содержит консервативный остаток лизина, необходимый для протеолиза.
Хотя в норме протеасома высвобождает очень короткие пептиды, иногда продукты протеасомного разрушения сами по себе являются биологически активными функциональными молекулами. Некоторые транскрипционные факторы, в числе которых компонент комплекса NF-κB млекопитающих, синтезируются в виде неактивных предшественников, которые становятся активными после убиквитинирования и протеасомной деградации. Для подобной активации разрыв пептидных связей должен происходить не на концах молекулы, а в её средней части. Было высказано предположение, что собственно субстратом для протеасомы являются длинные петли этих белков, при этом большая часть белковой молекулы не попадает внутрь протеасомы. Похожий механизм активации был выявлен и у дрожжей. Он получил название убиквитин-протеасомо-зависимого процессинга.
Убиквитин-независимое разрушение
Хотя в большинстве случаев субстраты протеасом должны быть полиубиквитинированы, из этого правила известно несколько исключений, в частности, в тех случаях, когда протеасома участвует в нормальном посттрансляционном процессинге белка. Субъединица комплекса NF-κB млекопитающих p105 должна быть разрушена до p50, который и входит в состав активного комплекса. Некоторые нестабильные белки, содержащие длинные неуложенные участки, также, вероятно, разрушаются протеасомами без убиквитиновых цепочек. Наиболее изученным убиквитин-независимым субстратом протеасомы является орнитиндекарбоксилаза. Убиквитин-независимому разрушению могут подвергаться некоторые регуляторы клеточного цикла. Наконец, белки с ненормальной структурой или сильно окисленные белки в условиях клеточного стресса разрушаются протеасомами независимо от частицы 19S и убиквитина.
Эволюция

20S-протеасома есть у всех эукариот и необходима для жизнедеятельности эукариотической клетки. У ряда прокариот, в числе которых многие археи и бактерии порядка Actinomycetales, имеются гомологи 20S-протеасомы. У большинства же бактерий есть гены теплового шока hslV и hslU, белковые продукты которых формируют мультимерную протеазу, состоящую из двух колец. Было высказано предположение, что белок hslV, возможно, похож на предка 20S-протеасомы. Как правило, hslV не является строго необходимым для бактериальной клетки и обнаруживается не у всех бактерий, однако у некоторых протистов есть и 20S-протеасома, и hslV. У многих бактерий выявляются другие гомологи протеасомы и связанной АТФазы, например, ClpP и ClpX. Многообразием гомологов протеасомы можно объяснить то, что система HslUV не является строго необходимой для бактериальных клеток.
Анализ последовательностей показал, что каталитические β-субъединицы в ходе эволюции обособились раньше, чем α-субъединицы, играющие преимущественно структурную роль. У бактерий, имеющих 20S-протеасому, последовательности β-субъединиц очень похожи на таковые у архей и эукариот, а последовательности α-субъединиц схожи в гораздо меньшей степени. Бактерии могли получить 20S-протеасому посредством горизонтального переноса генов, а диверсификация субъединиц протеасомы у эукариот есть следствие множественных дупликаций генов.
Клеточные функции
Клеточный цикл находится под контролем циклинзависимых киназ (CDK), которые активируются белками циклинами. Митотические циклины существуют всего несколько минут и являются одними из самых короткоживущих клеточных белков. После того как комплекс циклина с CDK выполнит свою функцию, циклин полиубиквитинируется и разрушается протеасомой, из-за чего соответствующая ему CDK становится неактивной, и наступает следующая фаза клеточного цикла. В частности, для выхода из митоза необходимо протеасомное разрушение циклина B. При прохождении через контрольную точку клеточного цикла, известную как точка рестрикции и находящуюся между G1-фазой и S-фазой, происходит протеасомное разрушение циклина А, причём его убиквитинирование осуществляет комплекс стимуляции анафазы (APC), который представляет собой Е3-убиквитинлигазу. APC и комплекс SCF — два ключевых фактора деградации циклинов. Более того, сам комплекс SCF регулируется APC через убиквитинирование адаптерного белка Skp2, который подавляет активность SCF до перехода из G1-фазы в S-фазу.
Отдельные компоненты 19S-частицы имеют свои собственные клеточные функции. Так, один из компонентов 19S-частицы, известный как ганкирин, является онкопротеином, который прочно связывается с циклинзависимой киназой 4 (CDK4) и, взаимодействуя с убиквитинлигазой MDM2, играет важнейшую роль в распознавании убиквитинированного p53. Ганкирин подавляет апоптоз и сверхэкспрессируется в некоторых видах раковых опухолей, например, гепатоцеллюлярной карциноме.
У растений фитогормоны ауксины стимулируют протеасомное разрушение Aux/IAA — репрессоров транскрипционных факторов. Эти белки убиквитинируются SCFTIR1 — комплексом SCF с рецептором ауксина TIR1. В результате разрушения Aux/IAA происходит дерепрессия транскрипционных факторов семейства факторов ответа на ауксин (ARF), из-за чего активируется экспрессия подконтрольных им генов. Клеточные эффекты активации ARF зависят от стадии развития растения, однако чаще всего они регулируют направление роста корней и жилок листа. Специфичность ответа на дерепрессию ARF, вероятно, обеспечивает чёткое соответствие между определёнными белками семейств Aux/IAA и ARF.
Протеасомы играют важную роль в протекании апоптоза, стимулируя убиквитинирование белков, хотя ведущая роль в разрушении белков при апоптозе принадлежит каспазам. При апоптозе протеасомы, находящиеся в ядре погибающей клетки, перемещаются в состав так называемых блебов, отшнуровывающихся от клеточной мембраны (блеббинг мембраны является характерной чертой апоптоза). В клетках разных видов ингибирование протеасом оказывает разное воздействие на апоптоз. В большинстве случаев протеасомы не строго необходимы для апоптоза, хотя у большинства клеток ингибирование протеасом запускает апоптоз. Важную роль в запуске апоптоза играет нарушение работы слаженной системы деградации белков, стимулирующих пролиферацию и деление клеток. Однако клетки некоторых типов, например, дифференцированные клетки в G0-фазе, такие как тимоциты и нейроны, под действием ингибиторов протеасом не уходят в апоптоз. Механизм этого эффекта не ясен, но, вероятно, специфичен для покоящихся клеток или обусловлен дифференциальной активностью проапоптотической киназы JNK. Способность ингибиторов протеасом запускать апоптоз в быстро делящихся клетках используется в некоторых недавно разработанных препаратах противораковой химиотерапии, таких как бортезомиб и салиноспорамид А.
В условиях клеточного стресса, таких как инфекция, тепловой шок, окислительные повреждения, экспрессируются белки теплового шока, которые распознают неправильно уложенные или денатурированные белки и направляют их на протеасомное разрушение. Было показано, что шапероны Hsp27 и Hsp90 задействованы в повышении активности убиквитин-протеасомной системы, хотя они не принимают непосредственного участия в этом процессе. Ещё один шаперон, Hsp70, связывается с выставленными наружу гидрофобными участками неуложенных белков (в норме такие участки обращены внутрь молекулы) и привлекает убиквитинлигазы вроде CHIP, которые направляют белки на деградацию в протеасомах. Схожие механизмы направляют на разрушение окисленные белки. Например, ядерные протеасомы регулируются поли(АДФ-рибоза)-полимеразами (PARP) и активно разрушают окисленные гистоны. Окисленные белки часто формируют большие аморфные скопления внутри клетки, и 20S-частица способна разрушать их без 19S-частицы, независимо от гидролиза АТФ и убиквитина. Однако сильные окислительные повреждения повышают риск перекрёстного соединения фрагментов белков, из-за чего они становятся устойчивыми к протеолизу. Большие по размеру и количеству скопления окисленных белков связаны со старением.
Роль в иммунной системе
Протеасомы играют важнейшую роль в функционировании адаптивного иммунитета. В протеасомах антигенпрезентирующих клеток белки вторгнувшегося в организм патогена разрушаются до пептидов, которые экспонируются наружу молекулами главного комплекса гистосовместимости I класса (MHCI). В этом процессе могут принимать участие как обычные, постоянно экспрессирующиеся протеасомы, так и специализированные иммунопротеасомы. Их экспрессия запускается интерфероном γ, и образуемые ими пептиды имеют идеальные размер и состав для экспонирования MHC. Во время иммунного ответа повышается экспрессия регуляторной субъединицы 11S, которая регулирует образование лигандов MHC, а также специализированных β-субъединиц β1i, β2i и β5i, имеющих слегка изменённую субстратную специфичность. Иммунопротеасомы — это протеасомы, содержащие такие специализированные β-субъединицы. В тимусе экспрессируется другой вариант субъединицы β5i — β5t, что приводит к образованию специфичных для тимуса тимопротеасом, функции которых неясны.
Сила связывания лиганда MHCI зависит от аминокислотного состава C-конца белка-лиганда, так как именно его C-конец посредством водородных связей связывается с особым участком на поверхности MHCI, который называется B-карманом. Многие аллели MHCI наиболее хорошо связываются с гидрофобными C-концами, и пептиды, образуемые иммунопротеасомами, как правило, имеют гидрофобные C-концы.
Так как протеасомы задействованы в активации NF-κB — антиапоптотического и провоспалительного регулятора экспрессии цитокинов, они играют роль в развитии воспалительных и аутоиммунных заболеваний. Повышенный уровень экспрессии протеасом связан с выраженностью заболевания и наблюдается при таких аутоиммунных заболеваниях, как системная красная волчанка и ревматоидный артрит.
Протеасомы принимают участие во внутриклеточном протеолизе, опосредованном антителами, которому подвергаются связанные с антителами вирусные частицы (вирионы). Белок TRIM21 связывается с иммуноглобулином G и направляет вирион на протеасомное разрушение.
Ингибиторы протеасом
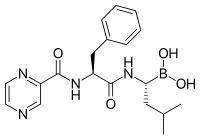
Ингибиторы протеасом демонстрируют выраженную противоопухолевую активность в культурах клеток, вызывая апоптоз нарушением деградации белков. Благодаря селективному проапоптотическому действию на раковые клетки ингибиторы протеасом прошли успешные клинические испытания на животных и людях.
Первым идентифицированным ингибитором протеасом непептидной природы стал лактацистин, синтезируемый бактериями рода Streptomyces. Лицензией на лактацистин обладает компания Takeda Pharmaceutical. Он нашёл широкое применение в исследовательской работе в области биохимии и клеточной биологии. Лактацистин ковалентно модифицирует N-концевые остатки треонина β-субъединиц, особенно субъединицы β5, которая обладает химотрипсин-подобной активностью. Благодаря лактацистину удалось установить, что протеасома является аминоконцевой треониновой протеазой (первой представительницей нового класса протеаз).
Бортезомиб (торговое название Велкад), разработанный компанией Millennium Pharmaceuticals, стал первым ингибитором протеасом, который начал использоваться в противораковой химиотерапии. Его используют для лечения множественной миеломы. При множественной миеломе в плазме крови выявляется высокий уровень пептидов протеасомного происхождения, который снижается до нормального при лечении бортезомибом. Исследования на животных показали, что бортезомиб может быть эффективен при раке поджелудочной железы. С начала XXI века проводятся доклинические и клинические испытания эффективности бортезомиба при лечении других видов рака B-клеточного происхождения, в частности, некоторых неходжкинских лимфом. Клинические испытания показали эффективность бортезомиба в сочетании со стандартной химиотерапией в борьбе с B-клеточной острой лимфобластной лейкемией. Ингибиторы протеасом в условиях культуры клеток убивают клетки некоторых лейкемий, которые устойчивы к глюкокортикоидам.
Препарат ритонавир (торговое название Норвир) был разработан как ингибитор протеаз для лечения ВИЧ-инфекции. Однако оказалось, что он подавляет не только свободные протеазы, но и протеасомы — точнее, блокирует химотрипсиноподобную активность протеасомы, при этом немного повышая трипсиноподобную активность. Исследования на животных показали, что ритонавир может подавлять рост клеток глиомы.
Как показали эксперименты на животных моделях, ингибиторы протеасом могут быть эффективны при лечении аутоиммунных расстройств. Изучение мышей с пересаженной человеческой кожей показало, что под действием ингибиторов протеаз размер язв, вызванных псориазом, уменьшался. Ингибиторы протеаз также оказались эффективны против астмы на животных моделях.
Мечение и подавление протеасом имеет важное значение для изучения работы протеасом как in vitro, так и in vivo. Чаще всего в исследовательской практике применяют такие ингибиторы протеасом, как лактацистин и пептидный альдегид MG132. Для мечения активных сайтов протеасом были разработаны специфические флуоресцентные ингибиторы.
Клиническое значение
Протеасомы и их субъединицы важны для медицины не только как молекулярная основа многих заболеваний, но и как перспективная мишень для многих лекарственных препаратов. Возможно, протеасомы могут быть использованы в качестве биомаркеров (в частности, биомаркеров некоторых аутоиммунных заболеваний). Нарушения в работе протеасом были выявлены при нейродегенеративных, сердечно-сосудистых, воспалительных и аутоиммунных заболеваниях, а также многих видах рака. Также они могут быть связаны с опухолями мозга, например, астроцитомами.
Несколько экспериментальных и клинических исследований указали на связь дисфункции протеасом с многими нейро- и миодегенеративными заболеваниями, в числе которых болезнь Альцгеймера, болезнь Паркинсона, болезнь Пика, боковой амиотрофический склероз и другие заболевания моторных нейронов, болезнь Гентингтона, болезнь Крейтцфельдта — Якоба, несколько редких нейродегенеративных заболеваний, связанных с деменцией, полиглутаминовые заболевания, мышечные дистрофии и миопатия телец включения. При дисфункции протеасом в нервной ткани образуются большие нерастворимые скопления неуложенных белков, что часто наблюдается при нейродегенеративных заболеваниях (например, при болезни Паркинсона образуются так называемые тельца Леви). Однако молекулярные основы нейротоксичности белковых скоплений неясны. Исследования на дрожжах показали, что клетки наиболее чувствительны к токсическому действию α-синуклеина (главного белка телец Леви) в условиях ингибирования протеасом. Неправильная работа протеасом может лежать в основе таких когнитивных проблем, как расстройства аутистического спектра.
Нарушения, касающиеся функционирования протеасом, связаны с ишемической болезнью сердца, гипертрофией желудочков и инфарктом миокарда. Так как протеасомы связаны с ответом клеток на стимулирующие сигналы, их дисфункции могут приводить к раку. Под контролем протеасом находится численность множества белков, связанных с развитием рака: p53, c-Jun, c-Fos, NF-κB, c-Myc, HIF-1α, MATα2, STAT3 и другие. Протеасомы разрушают многие белки, функционирующие как супрессоры опухолей, например, adenomatous polyposis coli и VHL, а также некоторые протоонкогены (Raf, Myc, Myb, Rel, Src, Mos, Abl). Регулируя активацию NF-κB, который активирует экспрессию провоспалительных цитокинов, простагландинов и оксида азота (NO), протеасомы вовлечены в регуляцию воспаления. Влияя на разрушение циклинов и ингибиторов циклин-зависимых киназ, протеасомы выступают как регуляторы пролиферации лейкоцитов при воспалении.
Ссылки
| В библиографических каталогах |
|---|
|
Эндомембранная система |
|
|---|---|
| Цитоскелет | |
| Эндосимбионты | |
| Другие внутренние органеллы | |
| Внешние органеллы | |