
| Парафилетическая группа организмов | |||||||
|---|---|---|---|---|---|---|---|
 | |||||||
| Название | |||||||
| Прокариоты | |||||||
| Статус названия | |||||||
| Устаревшее таксономическое | |||||||
| Научное название | |||||||
| Procaryota | |||||||
| Родительский таксон | |||||||
| Наддомен Биота | |||||||
| Представители | |||||||
| Кладистически вложенные, но традиционно исключаемые таксоны | |||||||
| |||||||

Прокарио́ты (лат. Procaryota, от др.-греч. πρό ‘перед’ и κάρυον ‘ядро’), или доя́дерные — одноклеточные живые организмы, не обладающие (в отличие от эукариот) оформленным клеточным ядром и другими внутренними мембранными органеллами (такими как митохондрии или эндоплазматический ретикулум, за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий).
Прокариоты не развиваются и не дифференцируются в многоклеточную форму. Некоторые бактерии растут в виде волокон или клеточных масс, но каждая клетка в колонии одинакова и способна к самостоятельной жизни.
С точки зрения биомассы и количества видов, прокариоты являются наиболее представительной формой жизни на Земле. Например, прокариоты в море составляют 90 % от общего веса всех организмов, в одном грамме плодородной почвы более 10 миллиардов бактериальных клеток. Известно около 3000 видов бактерий и архей, но это число, вероятно, составляет менее 1 % от всех существующих видов в природе.
Классификация
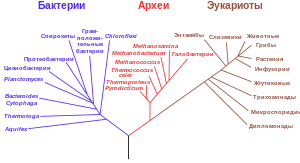
Название «прокариоты» (prokaryotes) было предложено Эдуардом Шаттоном в 1925 году. Однако в таксономическом смысле Шаттон не определил этот термин, то есть он не ставил таксономический диагноз. Несмотря на это, в биологических классификациях предлагаемое разделение организмов на прокариотические и эукариотические оставалось до 1990-х годов.
Большинство прокариотических клеток очень малы по сравнению с эукариотическими клетками. Типичная бактериальная клетка имеет размер около 1 мкм, тогда как эукариотические клетки имеют большой размер от 10 до 100 мкм. Типичная прокариотическая клетка примерно такого же размера, как эукариотическая митохондрия.
Этот набор характеристик отличает их от эукариот (ядерных организмов), которые имеют клеточные ядра и могут быть как одноклеточными, так и многоклеточными. Разница между структурой прокариот и эукариот крупнейшая среди групп организмов. Большинство прокариот — бактерии, и эти два термина ранее рассматривались как синонимы. Однако, американский ученый Карл Вёзе предложил разделение прокариот на бактерий и архей (Bacteria и Archaea, сначала Eubacteria и Archaebacteria) через существенные генетические различия между этими группами. Система разделения на эукариот, бактерий и архей сейчас считается признанной и называется Системой трех доменов.
На протяжении большей части XX века прокариоты считались единой группой и классифицировались по биохимическим, морфологическим и метаболическим особенностям. К примеру, микробиологи пытались классифицировать микроорганизмы в зависимости от формы клеток, деталей строения клеточной стенки и потребляемых микроорганизмами веществ. В 1965 году было предложено устанавливать степень родства разных прокариот на основании сходства строения их генов. Этот подход, филогенетика, в наши дни является основным. В конце XX-го века молекулярные исследования дали ключевую информацию для понимания эволюционного прошлого прокариот и доказали парафилетический характер этой группы организмов. Оказалось, что археи, обнаруженные в 1970-х годах, так же далеки от бактерий, как и от эукариот, а в некоторых отношениях даже ближе к последним (см. Интрон).
| Свойственно археям и бактериям | Свойственно археям и эукариотам | Свойственно только археям |
|---|---|---|
| Нет оформленного ядра и мембранных органелл | Нет пептидогликана (муреина) | Структура клеточной стенки (к примеру, клеточные стенки некоторых архей содержат псевдомуреин) |
| Кольцевая хромосома | ДНК связана с гистонами | В клеточной мембране присутствуют липиды, содержащие простую эфирную связь |
| Гены объединены в опероны | Трансляция начинается с метионина | Структура флагеллинов |
| Схожие РНК-полимераза, промоторы и другие компоненты транскрипционного комплекса, есть интроны и процессинг РНК | Структура рибосом (некоторые признаки сближают с бактериями, некоторые — с эукариотами) | |
| Полицистронная мРНК | Схожие репликация и репарация ДНК | Структура и метаболизм тРНК |
| Размер клеток на несколько порядков меньше, чем у эукариот | Схожая АТФаза (тип V) |
Первоначально бактерии и цианобактерии, считающиеся отдельными группами, были объединены под названием прокариот (или царство Дробянки (лат. Monera)). Затем цианобактерии считались группой бактерий, а другая ветвь бактерий стала идентифицироваться как так называемые архебактерии (ныне археи).
Однако помимо получившей широкое признание системы Вёзе, все же существуют альтернативные системы групп высшего уровня.

Система «с двумя империями» (система с двумя надцарствами) была системой биологической классификации высшего уровня общего пользования до создания системы с тремя доменами. Онa классифицировалa жизнь разделяя ее на Прокариот и Эукариот. Когда была введена система с тремя доменами, некоторые биологи все же предпочли систему с двумя империями, утверждая, что система с тремя доменами переоценила разделение между археями и бактериями. Однако, учитывая текущее состояние знаний и быстрый прогресс в области биологии, особенно благодаря генетическому анализу, эта точка зрения практически исчезла.

Клада Neomura состоящая из двух доменов: Archaea и Eukaryota. была предложена английским биологом Томасом Кавалир-Смитом, теория предполагает, что группа эволюционировала от Bacteria, и одним из важнейших изменений была замена пептидогликана клеточной стенки другими гликопротеинами, происхождение представителей наддомена от грамположительных бактерий (Firmicutes и Actinobacteria) подтверждается также по результатам сравнительного анализа генов белков семейства HSP90. В мае 2015 года опубликованы результаты исследования, предлагающего выделение нового типа архей — Lokiarchaeota с предполагаемым родом Lokiarchaeum. Он был выделен на основании генома, собранного при метагеномном анализе образцов, полученных рядом с гидротермальными источниками в Атлантическом океане на глубине 2,35 км. Филогенетический анализ показал, что Lokiarchaeota и эукариоты образуют монофилетический таксон. Геном Lokiarchaeota содержит около 5400 генов, кодирующих белки. Среди них были обнаружены гены, близкие к генам эукариот. В частности, гены, кодирующие белки, отвечающие за изменение формы клеточной мембраны, определение формы клетки и динамический цитоскелет. Результаты этого исследования служат подтверждением так называемой двухдоменной, или эоцитной гипотезы, согласно которой эукариоты появились как особая группа внутри архей, близкая к Lokiarchaeota и приобретшая митохондрии в результате эндосимбиоза. Исторически различают пять царств живых организмов: животные, растения, грибы, бактерии и вирусы. С 1977 года добавлены царства протисты и археи, с 1998 года — хромисты.
Подразделения надцарства «Прокариоты»
Традиционно таксономия бактерий была основана на сходствах и различиях в морфологии (таксономия Линнея). До появления молекулярной филогении, надцарство Monera (как в то время были известны домены бактерии и археи) былo разделенo на четыре филa. Грамo-результаты были наиболее часто использ. в качестве инструмента классификации, В 1962 году Р. Г. Мюррей предложил разделить бактерии на 4 таксона (типa) на основе типов клеточных стенок:
- Грамотрицательные грацилюты (Gracilicutes) с тонкой клеточной стенкой и небольшим количеством пептидогликана
- Грамположительные «Фирмакуты» («Firmacutes»), с более толстой клеточной стенкой и большим количеством пептидогликана (название было позже изменено в «Firmicutes»), и сегодня он включает в себя Молликуты (Mollicutes), но исключает актинобактерии);
- «Молликуты» (Микоплазмы), без клеточной стенки (грамм-переменная, переименована в Тенерикуты (Tenericutes).)
- Мендосикуты (неравномерное окрашивание по Граму, «метаногенные бактерии», теперь известные как метаногены и классифицируемые как археи): бактерии с клеточной стенкой без пептидогликана.
Эта система классификации была заброшена в пользу трехдоменной системы, основанной на молекулярной филогении, начатой C. Woese.
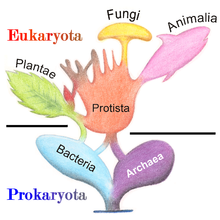
Древо жизни
Все царства объединяют в четыре надцарства, или домена: бактерии, археи, эукариоты и вирусы. К домену бактерии относят царство бактерии, к домену археи — царство археи, к домену вирусы — царство вирусы (для простоты понимания в таблице есть сразу разделение по Балтимору на 7 классов, так бы было одно царство вирусов, один отдел вирусов, только потом 7 классов), к домену эукариоты — все остальные царства. Нужно также понимать, что вирусы не могут жить сами по себе, поэтому в системе Вёзе они не указываются.
| Биота |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||
Описание
Для клеток прокариот характерно отсутствие ядерной оболочки, ДНК упакована без участия гистонов. Тип питания осмотрофный и автотрофный (фотосинтез и хемосинтез).
Единственная крупная кольцевая (у некоторых видов — линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид), не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (синезелёные водоросли), и археи. Потомками прокариотических клеток являются органеллы эукариотических клеток — митохондрии и пластиды.
Прокариоты разделяют на два таксона в ранге домена (надцарства):
Изучение бактерий привело к открытию горизонтального переноса генов, который был описан в Японии в 1959 году. Этот процесс широко распространён среди прокариот, а также у некоторых эукариот.
Открытие горизонтального переноса генов у прокариот заставило по-другому взглянуть на эволюцию жизни. Ранее эволюционная теория базировалась на том, что виды не могут обмениваться наследственной информацией.
Прокариоты могут обмениваться генами между собой непосредственно (конъюгация, трансформация) а также с помощью вирусов — бактериофагов (трансдукция).
Прокариоты имеют прокариотический цитоскелет, хотя и более примитивный, чем у эукариот. Помимо гомологов актина и тубулина (MreB и FtsZ), спирально расположенный строительный блок жгутика, флагеллин является одним из наиболее значимых цитоскелетных белков бактерий, поскольку он обеспечивает структурные предпосылки хемотаксиса, основного клеточного физиологического ответа бактерий на химический раздражитель. По крайней мере, некоторые прокариоты также содержат внутриклеточные структуры, которые можно рассматривать как примитивные органеллы. Мембранные органеллы (или внутриклеточные мембраны) известны у некоторых групп прокариот, такиe как вакуоли или мембранные системы, которые обладают особыми метаболическими свойствами, такими как фотосинтез или хемолитотрофия. Кроме того, некоторые виды также содержат заключенные в углеводы микрокомпартменты, которые выполняют различные физиологические функции (например, карбоксисомы или газовые вакуоли).

У прокариот большая часть химических реакций метаболизма происходит в цитозоле, и лишь небольшая их часть происходит в мембранах и периплазматическом пространстве.
До 1990-х годов считалось, что опероны встречаются только у прокариот, но с момента открытия первых оперонов у эукариот появилось больше доказательств того, что они встречаются чаще, чем предполагалось ранее. В целом, экспрессия прокариотических оперонов приводит к образованию полицистронных мРНК, в то время как эукариотические опероны приводят к моноцистронным мРНК.
Литотрофы могут образовывать симбиотические отношения, и в этом случае они называются «прокариотическими симбионтами». Примером таких отношений может служить симбиоз хемолитотрофных бактерий с гигантскими многощетинковыми червями.
Характерные особенности

Характерные особенности прокариотов:
- отсутствие чётко оформленного ядра;
- наличие жгутиков, плазмид и газовых вакуолей;
- наличие структур, в которых происходит фотосинтез;
- формы размножения: бесполый способ, имеется псевдополовой процесс (в результате которого происходит лишь обмен генетической информацией, без увеличения числа клеток);
- размер рибосомы — 70S (по коэффициенту седиментации различают и рибосомы других типов, а также субчастицы и биополимеры, входящие в состав рибосом);
- молекула ДНК уложена в виде петли, комплексируется некоторыми гистоновыми белками, образуя нуклеоид. Основная масса ДНК (95 %) активно транскрибируется в каждый данный момент времени.
Классификация организмов по типу метаболизма
Все живые организмы можно разделить на восемь основных групп в зависимости от используемого: источника энергии, источника углерода и донора электронов (оксисляемого субстрата).
- В качестве источника энергии живые организмы могут использовать: энергию света (фото-) или энергию химических связей (хемо-). Дополнительно для описания паразитических организмов использующих энергетические ресурсы хозяйской клетки применяют термин паратроф.
- В качестве донора электронов (восстановителя) живые организмы могут использовать: неорганические вещества (лито-) или органические вещества (органо-).
- В качестве источника углерода живые организмы используют: углекислый газ (авто-) или органические вещества (гетеро-). Иногда термины авто- и гетеротроф используют в отношении других элементов, которые входят в состав биологических молекул в восстановленной форме (например азота, серы). В таком случае «автотрофными по азоту» организмами являются виды, использующие в качестве источника азота окисленные неорганические соединения (например, растения; могут осуществлять восстановление нитратов). А «гетеротрофными по азоту» являются организмы не способные осуществлять восстановление оксисленных форм азота и использующие в качестве его источника органические соединения (например, животные, для которых источником азота служат аминокислоты).
Название типа метаболизма формируется путём сложения соответствующих корней и добавлением в конце корня -троф-. В таблице представлены возможные типы метаболизма с примерами:
| Источник энергии |
Донор электронов | Источник углерода | Тип метаболизма | Примеры |
|---|---|---|---|---|
| Солнечный свет Фото- |
Органические вещества -органо- |
Органические вещества -гетеротроф |
Фотоорганогетеротрофы | Пурпурные несерные бактерии, Галобактерии, Некоторые цианобактерии. |
| Углекислый газ -автотроф |
Фотоорганоавтотрофы | Редкий тип метаболизма, связанный с окислением неусваиваемых веществ. Характерен для некоторых пурпурных бактерий. | ||
| Неорганические вещества -лито-* |
Органические вещества -гетеротроф |
Фотолитогетеротрофы | Некоторые цианобактерии, пурпурные и зелёные бактерии, также гелиобактерии. | |
| Углекислый газ -автотроф |
Фотолитоавтотрофы | Высшие растения, Водоросли, Цианобактерии, Пурпурные серные бактерии, Зелёные бактерии. | ||
| Энергия химических связей Хемо- |
Органические вещества -органо- |
Органические вещества -гетеротроф |
Хемоорганогетеротрофы | Животные, Грибы, Большинство микроорганизмов редуцентов. |
| Углекислый газ -автотроф |
Хемоорганоавтотрофы | Окисление трудноусваиваемых веществ, например факультативные метилотрофы, окисляющие муравьиную кислоту. | ||
| Неорганические вещества -лито-* |
Органические вещества -гетеротроф |
Хемолитогетеротрофы | Метанобразующие археи, Водородные бактерии. | |
| Углекислый газ -автотроф |
Хемолитоавтотрофы | Железобактерии, Водородные бактерии, Нитрифицирующие бактерии, Серобактерии. |
- Некоторые авторы используют -гидро- когда в качестве донора электронов выступает вода.
Метаболические возможности прокариот значительно разнообразнее по сравнению с эукариотами, которые характеризуются фотолитоавтотрофным и хемоорганогетеротрофным типом метаболизма.
Некоторые виды микроорганизмов могут в зависимости от условий среды (освещение, доступность органических веществ и т. д.) и физиологического состояния осуществлять метаболизм разного типа. Такое сочетание нескольких типов метаболизма описывается как миксотрофия.
Морфология и размеры

Прокариотические клетки имеют различные формы; четыре основные формы бактерий:
- Кокки — сферическая
- Бациллы — палочковидные
- Спирохета — спиралевидная
- Вибрионы — по форме напоминает запятую
- Археон Haloquadratum имеет плоские квадратные клетки.
Большинство прокариот имеют размер от 1 до 10 мкм, но их размер может варьироваться от 0,2 мкм (Mycoplasma genitalium) до 750 мкм (Thiomargarita namibiensis). С диаметром клетки около 0,12—0,20 мкм Candidatus Pelagibacter ubique является одним из мельчайших известных самореплицирующихся организмов. Прокариотические клетки обычно намного меньше, чем эукариотические клетки. Следовательно, прокариоты имеют большее отношение площади поверхности к объему, что обеспечивает им более высокую скорость метаболизма, более высокую скорость роста и, как следствие, более короткое время генерации, чем у эукариот. Минимальный размеры генома прокариот имеют определенные теоретические ограничения. В 1996 году Аркадий Мушегян и Евгений Кунин (Национальный центр биотехнологической информации, США) предположили, что 256 ортологичных генов, общих для грамотрицательной бактерии Haemophilus influenzae и грамположительной Mycoplasma genitalium, являются хорошим приближением к минимальному набору генов бактериальной клетки. В 2004 году группа исследователей из университета Валенсии (Испания) предложила набор из 206 кодирующих белки генов, полученный в результате анализа нескольких бактериальных геномов.
Учёные из группы Крейга Вентера занимались созданием организма с минимальным искусственно синтезированным геномом, начиная с 1995 года. В 1995 году они секвенировали геном возбудителя заболеваний мочеполовой системы человека Mycoplasma genitalium — минимальный среди известных к настоящему времени организмов, способных к самовоспроизведению. Этот микроорганизм содержит 517 генов, из которых 482 кодируют белки. Полный объём генома составляет 580 тыс. нуклеотидных пар. К 1999 году, анализируя расположение транспозонов в секвенированных геномах, удалось установить, что жизненно необходимыми для организма являются от 265 до 350 генов и более 100 генов имеют неизвестное назначение. Дальнейшие исследования к 2005 году расширили список жизненно необходимых генов. Команда учёных систематически удаляла гены, чтобы найти минимальный набор генов, необходимый для жизни. Результат: 382 гена. Эта работа была также известна как Проект Минимального Генома.
В дальнейшем были обнаружены прокариотические геномы ещё меньшего размера, но все они принадлежат облигатным симбионтам — не способным к автономному существованию организмам.
В 2003 году был секвенирован геном Nanoarchaeum equitans размером 490 885 пар. Установлено также, что несеквенированный геном вида Buchnera имеет длину около 450 тыс. пар.
Наименьший из расшифрованных к настоящему моменту бактериальных геномов — геном внутриклеточного эндосимбионта листоблошек бактерии Carsonella, состоящий из 159 662 нуклеотидных пар и содержащий всего 182 гена, кодирующих белки. Этот геном был секвенирован японскими исследователями в 2006 году. В 2009 году был проведён анализ некультивируемых клеток ARMAN из рудниковой биоплёнки с использованием трёхмерной криоэлектронной томографии. Оказалось, что размер клеток ARMAN соответствует нижнему предсказанному пределу для живых клеток: их объём составляет 0,009—0,04 мкм³. Выяснилось также, что в клетках ARMAN имеется в среднем 92 рибосомы на клетку, в то время как в клетке кишечной палочки Escherichia coli содержится около 10 тысяч рибосом. Судя по всему, ARMAN обходятся очень малым числом метаболитов на одну клетку, что поднимает вопрос о минимальных потребностях живых клеток. 3D-реконструкция клеток ARMAN в естественной среде обитания показала, что некоторые клетки ARMAN прикрепляются к другим археям из порядка Thermoplasmatales. Клетки этих архей проникают сквозь клеточную стенку ARMAN, достигая цитоплазмы. Природа этого взаимодействия неясна; возможно, здесь имеет место некоторая разновидность паразитизма или симбиоза. Возможно, ARMAN получают от других архей те метаболиты, которые не могут синтезировать сами.
Однако некоторые прокариоты не полагаются на свой малый размер и соответствующее такому эволюционному вектору упрощению своего генотипа. Например бактерия Desulforudis audaxviator, найденная в пробах воды, полученных на глубине 2,8 км под землей приблизительно четыре микрометра в длину, выживает уже в течение миллионов лет на химических источниках пищи, которые происходят из-за радиоактивного распада минералов в окружающей породе. Это делает ее единственным видом, который известен как один в своей экосистеме. Физиология, которая позволяет ему жить в этих экстремальных условиях, является данью его необычайно большому геному, состоящему из 2157 генов вместо 1500 у бактерий такого типа.
Согласно опубликованным данным, размер варьируется от 0,58 мегабазы (1 мегабаза (Мб) — один миллион пар оснований (п.о.)) во внутриклеточном патогене Mycoplasma genitalium, до более 10 Мб у нескольких видов цианобактерий, за исключением Bacillus megaterium, геном которого составляет 30 Мб. Второй самый маленький из когда-либо опубликованных геномов — это Buchnera sp. APS, эндосимбионт зерновой тли Acyrthosiphon pisum, размером 641 кб. Недавно исследовательская группа охарактеризовала шесть геномов, меньших, чем даже у Mycoplasma, наименьший из которых — у Buchnera sp. CCE, эндосимбионт тли Cinara cedri, размером 0,45 Мб. Как правило, большинство геномов имеют размер менее 5 Мб. Возникает вопрос есть ли связь между размером генома и количеством генов? Размер прокариотического гена однороден, около 900—1000 п.о. Следовательно, можно оценить плотность генов в каждом секвенированном геноме. Плотность генов является более или менее постоянной, как у бактерий, так и у архей. Мы можем заключить, что, по крайней мере у прокариот, геномы имеют большее количество генов, а также являются более сложными чем у эукариот. То есть количество генов отражает стиль жизни. Таким образом, более мелкие бактерии являются специалистами, такими как облигатные паразиты и эндосимбионты, а более крупные бактерии являются универсалами и могут даже иметь определенную степень развития, такую как споруляция (Процесс образования спор) у Bacillus.

Плазмиды (небольшие молекулы ДНК, физически отдельные от хромосом и способные реплицироваться автономно) как правило, встречаются у бактерий, но изредка встречаются также у архей и эукариот. Чаще всего плазмиды представляют собой двухцепочечные кольцевые молекулы. Несмотря на способность к размножению, плазмиды, как и вирусы, не рассматриваются в качестве живых организмов. Размеры плазмид варьируют от менее чем 1 тысячи до 400—600 тысяч пар оснований (п. о.). Некоторые плазмиды содержатся в клетке в количестве одной-двух копий, другие — в количестве нескольких десятков. Плазмиды разных классов могут сосуществовать в клетке. В природе плазмиды обычно содержат гены, повышающие приспособленность бактерий к окружающей среде (например, обеспечивают устойчивость к антибиотикам). Если самые маленькие плазмиды содержат менее 2 тысяч пар оснований, то так называемые мегаплазмиды включают сотни тысяч пар оснований (обычно до 600 тыс.). В этом случае уже сложно провести чёткую границу между мегаплазмидой и минихромосомой. Некоторые виды бактерий могут одновременно содержать множество различных плазмид, так что их суммарный генетический материал превосходит по размеру таковой у самой бактерии. Например, симбиотическая почвенная бактерия Sinorhizobium meliloti содержит 3 репликона размером 3,65, 1,68 и 1,35 миллиона п. о. (мегабаз) соответственно вдобавок к её собственной хромосоме (6,69 мегабазы).
Размножение
Размножение бактерий происходит в три этапа. Когда популяция бактерий попадает в очень питательную среду, которая обеспечивает рост, клетки должны сначала адаптироваться к новой среде. Первая стадия развития (фаза называется лаг-фазой) характеризуется медленным ростом, когда клетки сначала адаптируются и готовятся к быстрому росту. Следующим шагом является логарифмическая фаза или экспоненциальный рост, что означает, что когда число измеряется после равного интервала времени, бактерии начинают размножаться с тем же фактором или коэффициентом, который усиливается числом интервалов. Это происходит до тех пор, пока питательные вещества не иссякнут.
После этой фазы третья фаза, которая называется «фазой сна», происходит там, где бактерии не размножаются.
И наконец финальная фаза роста — фаза смерти, при которой запас питательных веществ исчерпывается и бактерии погибают. Многие обитающие в обедненных средах прокариоты выживают в условиях, схожих с перманентным анабиозом, тем самым экономя энергию, и размножаются невероятно медленно: раз в сотни или даже тысячи лет.
Продолжительность жизни
| Организм | Группа | Среда | Время удвоения, мин. |
|---|---|---|---|
| Escherichia coli | бактерии | глюкоза, соли | 17 |
| Bacillus megaterium | бактерии | сахароза, соли | 25 |
| Streptococcus lactis | бактерии | молоко | 26 |
| Staphylococcus aureus | бактерии | сердечный бульон | 27-30 |
| Lactobacillus acidophilus | бактерии | молоко | 66-87 |
| Myxococcus xanthus | бактерии | соли, экстракт дрожжей | 240 |
| Rhizobium japonicum | бактерии | манитол, соли, экстракт дрожжей | 344—461 |
| Mycobacterium tuberculosis | бактерии | синтетическая | 792—932 |
| Treponema pallidum | бактерии | яички кролика | 1980 |
Продолжительность жизни не имеет чёткого определения для одноклеточных организмов. Существует, тем не менее, несколько сроков, которые могут использоваться в таком качестве.
В первую очередь, при благоприятных условиях количество одноклеточных организмов экспоненциально возрастает, а характеристикой этого возрастания является время удвоения количества организмов или время одного поколения.
Другой характеристикой, аналогичной продолжительности жизни, являются характеристики процесса старения организмов. Одноклеточные организмы имеют два типа старения — «условное старение», или хронологическое старение в стационарной фазе, где возможно измерить среднюю или максимальную продолжительность жизни. Тем не менее, данные для сравнительной характеристики одноклеточных организмов отсутствуют. Другим типом старения является «репликативное старение», или старение материнской клетки во время каждого отделения от неё дочерней клетки, что обычно измеряется в количестве делений. Для дрожжей Saccharomyces cerevisiae максимальный репликативный возраст составляет около 25 делений, а для бактерии Caulobacter crescentis — около 130. Для остальных организмов данные отсутствуют.
Одноклеточные организмы отличаются высоким уровнем зависимости от условий окружающей среды. Со снижением температуры время удвоения и скорость старения снижаются практически для всех них. Многие одноклеточные организмы могут замедлить скорость роста в сотни раз, и сохраняться в замороженном виде на протяжении десятков лет и даже дольше. Также и наличие питательных веществ оказывает влияние на скорости роста и старения. Кроме того, многие одноклеточные организмы при неблагоприятных условиях формируют споры и другие неактивные формы, способные к существованию на протяжении многих лет.
Колонии
Обычно прокариотический организм представляет собой одну клетку. Иногда потомки нескольких отделений остаются связанными в колонии. В случае актиномицетов и многих цианобактерий «колония» представляет собой клеточную линию, между которой существует связь и даже определенное распределение функций. Истинная многоклеточность, однако, не встречается у прокариот. Одной из наиболее характерных черт прокариотической клетки является слабая компартментализация, то есть отсутствие множества внутренних секций, соединенных через систему элементарной мембраны. Для большинства прокариот цитоплазматическая мембрана является единственной мембранной системой клетки. Однако его топология часто сложна, так как мембранные складки проникают глубоко в цитоплазму. Цианобактерии являются единственным исключением из этого правила. В них аппарат фотосинтеза расположен на рядах герметичных мембранных мешков или тилакоидов, сходных по структуре и функции с тилакоидами хлоропластов. Однако в цианобактерии тилакоиды включены в определенные органеллы, но лежат непосредственно в цитоплазме.
История понятия
Монеры
Монеры — так Геккель назвал простейшие одноклеточные организмы без ядра. Так как присутствие ядра во многих случаях трудно констатируется, то первоначально, пока методы микроскопического исследования были сравнительно несовершенны, безъядерными считались очень многие формы жизни. Вопрос о монерах представляет некоторый интерес ввиду того, что первоначальное возникновение организмов на Земле, вероятно, произошло в форме тел, не дифференцированных ещё на ядро и протоплазму.
В настоящее время термин «монеры» не применяется.
Эволюция

Широко распространенная текущая модель эволюции первых живых организмов заключается в том, что это были некие формы прокариот, которые могли эволюционировать из протоклеток, в то время как эукариоты развивались позднее в истории жизни. Некоторые авторы подвергли сомнению этот вывод, утверждая, что текущий набор прокариотических видов, возможно, эволюционировал от более сложных эукариотических предков в процессе упрощения.

Другие утверждают, что три области жизни возникли одновременно, из набора разнообразных клеток, которые образовали единый генофонд. Это противоречие было обобщено в 2005 году:
Среди биологов нет единого мнения относительно положения эукариот в общей схеме эволюции клеток. Текущие мнения о происхождении и положении эукариот охватывают широкий спектр, включая мнения о том, что эукариоты возникли первыми в эволюции и что прокариоты происходят от них, что эукариоты возникли одновременно с эубактериями и археабактериями и, следовательно, представляют собой основную линию происхождения равного возраста и ранга, что и прокариоты, что эукариоты возникли в результате симбиотического события, влекущего за собой эндосимбиотическое происхождение ядра, что эукариоты возникли в результате симбиотического события, влекущего одновременное эндосимбиотическое происхождение жгутика и ядра, в дополнение ко многим другим модели, которые были рассмотрены и обобщены в другом месте.
Самые древние из известных окаменелых прокариот были заложены примерно 3,5 миллиарда лет назад, всего лишь около 1 миллиарда лет после образования земной коры. Эукариоты появляются только в окаменелостях позже и могут образоваться в результате эндосимбиоза нескольких предков прокариот. Самым древним известным ископаемым эукариотам около 1,7 миллиарда лет. Однако некоторые генетические данные свидетельствуют о том, что эукариоты появились еще 3 миллиарда лет назад.
Гипотеза вирусного происхождения эукариот предполагает, что эукариоты состоят из трёх предковых элементов: вирусный компонент, от которого произошло современное эукариотическое ядро; прокариотическая клетка, от которой эукариоты унаследовали цитоплазму и клеточную мембрану; а также ещё одна прокариотическая клетка, от которой произошли митохондрии и хлоропласты путём эндоцитоза. Возможно, клеточное ядро образовалось под воздействием нескольких заражений архейной клетки, уже содержащей бактерию — предшественника митохондрий, лизогенным вирусом.
В то время как Земля является единственным местом во вселенной, где, как известно, существует жизнь, некоторые полагают, что на Марсе имеются доказательства наличия ископаемых или живых прокариот. Однако эта возможность остается предметом значительных дебатов и скептицизма.
См. также
- Азотофиксация — Процесс восстановления молекулы азота и включения её в состав своей биомассы прокариотными микроорганизмами
- Сравнение строения клеток бактерий, растений, животных и грибов
- Эукариоты
- Систематика архей и Систематика бактерий
- Горизонтальный перенос генов
Литература
- Сергеев В. Н., Нолл Э.X., Заварзин Г. А. Первые три миллиарда лет жизни: от прокариот к эвкариотам // Природа. 1996. № 6. С. 54-68.
Ссылки
- Прокариоты на сайте «Ветеринарная медицина».
| Таксономия | ||||
|---|---|---|---|---|
|
| ||||
|
Классификация архей
| |
|---|---|
| Euryarchaeota | |
| |