
Конъюга́ция (от лат. conjugatio — соединение) — однонаправленный перенос части генетического материала (плазмид или бактериальной хромосомы) при непосредственном контакте двух бактериальных клеток. Открыт в 1946 году Джошуа Ледербергом и Эдвардом Татумом. Явление конъюгации было открыто и хорошо изучено у кишечной палочки (Escherichia coli), но в дальнейшем конъюгация была описана у множества как грамположительных, так и грамотрицательных бактерий. Посредством конъюгации бактерии обмениваются генетическим материалом, поддерживая своё генетическое разнообразие.
История изучения
Существование конъюгации у бактерий было показано в 1946 году Джошуа Ледербергом и Эдвардом Татумом. Они смешали два штамма кишечной палочки Escherichia coli, ауксотрофных (неспособных существовать на среде без определённого вещества) по разным веществам, инкубировали полученную культуру несколько часов в среде, содержащей все необходимые питательные вещества, и высеяли клетки на чашки с минимальной средой. Чтобы избежать искажения данных эксперимента реверсией некоторых мутаций, отвечающих за ауксотрофию, они использовали двойные и тройные ауксотрофы. Благодаря этому возможность появления клеток, не ауксотрофных по всем трём соединениям, за счёт реверсии мутаций была крайне низка. Когда на чашках появились колонии, исследователи заключили, что бактерии обменялись генами, которые восстанавливали нормальный фенотип. Однако Ледерберг и Татум не доказали, что ДНК передавалась между бактериями при непосредственном физическом контакте. Это удалось сделать Бернанду Дэвису в 1950 году. Он сконструировал U-образную трубку, разделённую напополам перегородкой, через которую могла проходить среда, но не бактериальные клетки. Далее Дэвис инкубировал в трубке несколько ауксотрофных штаммов, сильно перемешивая среду. После высеивания клеток на минимальную среду колоний не появилось. Таким образом, было показано, что для обмена ДНК, открытого Ледербергом и Татумом, был необходим физический контакт двух клеток. Впоследствии конъюгацию описали и у других бактерий, в том числе грамположительных, таких как Bacillus subtilis. Однако наиболее изучена конъюгация у E. coli, механизм которой детально рассматривается ниже.
Общий механизм
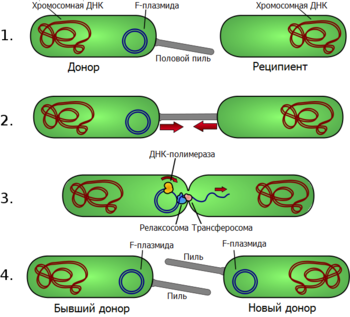
Под конъюгацией понимают перенос ДНК между бактериальными клетками при их непосредственном контакте. Как правило, при конъюгации передаются плазмиды, но у некоторых организмов передаваться может и хромосомная ДНК. При конъюгации имеет место однонаправленный перенос генетического материала от клетки-донора к клетке-реципиенту. Ниже рассматривается процесс конъюгации на примере E. coli, то есть у грамотрицательных бактерий; конъюгация у грамположительных бактерий рассматривается в разделе Конъюгация у других бактерий.
Образование пары
В подавляющем большинстве случаев конъюгация возможна лишь тогда, когда у донорской клетки есть плазмида, содержащая гены, обеспечивающие передачу ДНК. У E. coli и других грамотрицательных бактерий физический контакт между двумя клетками обеспечивается половыми пилями — полыми белковыми структурами на поверхности клеток. Структура половых пилей сильно варьирует: пили, кодируемые F-плазмидой, длинные, тонкие и гибкие, а пили, кодируемые плазмидой RP4, — короткие, толстые и жёсткие. Пили взаимодействуют с рецепторами на поверхности клетки-реципиента, за счёт чего конъюгирующие клетки образуют пару. Механизм образования пары клеток при конъюгации очень похож на работу бактериальной системы секреции IV типа, с помощью которой некоторые патогенные бактерии доставляют токсины непосредственно в клетку организма-хозяина.
Передача ДНК
Передача ДНК через контакт, образуемый пилем, начинается с того, что в специфическом участке передаваемой плазмиды, называемом ориджином переноса (oriT), особый белок вносит одноцепочечный разрыв (ник). Хеликаза, кодируемая передаваемой плазмидой, расплетает плазмидную ДНК, и разрезанная цепь переносится в клетку-реципиент своим 5'-концом вперёд. Параллельно с этим её 3'-конец удлиняется, восполняя уходящую цепь в плазмиде-доноре по механизму репликации типа катящегося кольца. Одноцепочечная ДНК, перенесённая в клетку-реципиент, также достраивается до двуцепочечной. Таким образом, конъюгация является репликативным процессом и, строго говоря, передаётся не сама плазмида, а её копия.
Мобилизация
К описанной выше передаче между клетками способны далеко не все плазмиды (их называют конъюгативными; например, таковой является F-плазмида E. coli). Иногда конъюгативная плазмида способствует передаче клетке-реципиенту неконъюгативной плазмиды при помощи процесса, который носит название мобилизации. В качестве примера можно рассмотреть мобилизацию неконъюгативной плазмиды ColE1. Она содержит ген mob, кодирующий специфическую нуклеазу, и сайт bom (аналогичный oriT), и белок Mob вносит одноцепочечный разрыв в плазмиду. По сути, для того, чтобы быть конъюгативной, плазмиде ColE1 недостаёт генов, которые кодировали бы аппарат переноса ДНК, поэтому она может передаваться в клетку-реципиент через аппарат передачи, кодируемый конъюгативной плазмидой. Иногда мобилизуемая плазмида может не иметь собственного гена mob, а одноцепочечный разрыв вносит в неё белок Mob близкородственной плазмиды.
Передача хромосомной ДНК
В подавляющем большинстве случаев при конъюгации передаётся плазмидная ДНК. Однако у некоторых бактерий возможна передача и хромосомной ДНК. Например, фрагменты геномной ДНК могут передаваться вместе с F-плазмидой у E. coli, и похожие системы есть у других бактерий, например, Pseudomonas aeruginosa. В некоторых случаях для передачи хромосомной ДНК необходима интеграция плазмиды в геномную ДНК бактерии, так что, по сути, геномная ДНК передаётся в составе плазмиды. Однако известны случаи, когда хромосомная ДНК передаётся в свободном виде, вероятно, по механизму, аналогичному мобилизации неконъюгативных плазмид.
При передаче плазмиды в ходе конъюгации клетка-реципиент получает полную одноцепочечную версию плазмиды, однако хромосомная ДНК не передаётся целиком в силу того, что её полная передача потребовала бы значительного времени. Так, для переноса полной хромосомы у E. coli потребовалось бы 100 минут (в то время как для передачи плазмиды длиной 40 тысяч пар оснований (п. о.) требуется 1 минута). Фрагмент геномной ДНК, перенесённый в клетку-реципиент, может использоваться последней для рекомбинации.
Конъюгация у E. coli
В 1952 году Уильям Хайес показал, что передача ДНК, описанная Ледербергом и Татумом, была полярна. Иными словами, в передаче участвовали клетка-донор (F+ от англ. fertility — «плодовитость») и клетка-реципиент (F-). Оказалось, что клетки F+-штамма содержат так называемую F-плазмиду. F-плазмида может находиться в клетке в двух состояниях: в свободном или в интегрированном в геном (эписомном).
При скрещивании F+-клеток и F--клеток клетка-донор не утрачивает свой F+-статус, но F--клетка при этом его приобретает. При таком скрещивании редко утрачивается ауксотрофия, поскольку клетка-реципиент получает лишь ДНК F-плазмиды без геномной ДНК.
Иная ситуация складывается, когда F--клетка конъюгирует с клеткой, у которой F-плазмида встроилась в геном. Такие клетки с встроенной в геном F-плазмидой называются Hfr от англ. high frequency of recombination, поскольку при их передаче клетка-реципиент часто получает и фрагмент геномной ДНК, который использует для рекомбинации. При интеграции F-плазмиды в геном все гены, необходимые для переноса ДНК, такие как гены, кодирующие белки пилей, остаются функциональными. В начале конъюгации между Hfr-клетами и F--клеткой в сайт oriT интегрированной F-плазмиды вносится одноцепочечный разрыв, и ДНК начинает переходить в клетку-реципиент, однако в неё сначала поступает лишь фрагмент F-хромосомы, а за ней — уже геномная ДНК. Как упоминалось выше, для переноса полного генома требуется около 100 минут, поэтому бактерии-партнёры разрывают контакт раньше, чем успеет перенестись вся геномная ДНК. Поскольку реципиент редко получает полную копию F-плазмиды, он остаётся F--клеткой.
Поскольку интегрированная в геном F-плазмида представляет собой эписому, она может покинуть бактериальный геном и восстановить свой автономный статус. Однако иногда при вырезании хромосомы происходит ошибка, и она захватывает с собой небольшой фрагмент геномной ДНК. Такую плазмиду называют F'-плазмидой. Поскольку F'-плазмида по-прежнему содержит все гены, необходимые для конъюгации, она остаётся конъюгативной и может передаваться F--клетке по механизму катящегося кольца. Однако по завершении конъюгации клетка-реципиент становится частично диплоидной (мерозиготой), потому что те участки генома, которые привнесла в неё F'-плазмида, содержатся в ней теперь в двойном комплекте. Конъюгацию, которую опосредует F'-плазмида, иногда называют сексдукцией. Стоит отметить, что между гомологичными участками на F'-плазмиде и хромосоме реципиента может происходить рекомбинация. Через F'-плазмиды бактериальные гены могут быстро распространяться в популяциях.
В процессе конъюгации задействовано множество белков. Одноцепочечный разрыв в области oriT и начальное расплетание цепей ДНК производит белковый комплекс, известный как релаксосома. У E. coli половой пиль, по сути, представляет собой систему секреции IV типа. Она состоит из множества копий белков Tra, в том числе TraA, которые формируют пиль, и TraD, которые обеспечивают процесс энергией за счёт гидролиза АТФ. Некоторые белки Tra находятся в клеточной мембране, некоторые продолжаются в периплазматическое пространство и проходят через слой пептидогликана, достигая внешней мембраны и её липополисахаридного слоя.
Регуляция

F-плазмида содержит особый регуляторный участок, в состав которого входит, в частности, ген traJ. Белок TraJ положительно регулирует экспрессию других генов tra, которые необходимы для образования и функционирования половых пилей. Антисмысловая РНК FinP и FinO образуют комплекс FinOP, который репрессирует образование белка TraJ и, таким образом, является ингибитором конъюгации. Описаны ещё 5 систем ингибирования переноса F-плазмиды и F-подобных плазмид в клетку-реципиент. Среди таких систем finW локализована в F-подобной плазмиде R455, системы finQ, finU и finV выявлены в ряде не-F-подобных плазмид, а система finC находится в многокопийном мутанте неконъюгативной плазмиды CloDF13. Все эти системы так или иначе регулируют гены tra. Сама F-плазмида тоже может ингибировать перенос других плазмид: так, она подавляет перенос плазмиды RP4 в 500—1000 раз.
Гены клетки-хозяина также могут влиять на конъюгативный перенос плазмид. Например, в геноме штамма K-12 E. coli выявлены гены sfrA и sfrB, которые регулируют некоторые гены tra. Кроме того, мутации в геномном гене fex понижают конъюгативный потенциал F-плазмиды.
Поверхностное исключение
Клетка, содержащая в том или ином виде конъюгативную плазмиду, не может выступать в роли реципиента при конъюгации. Это явление получило название поверхностного исключения и впервые было описано у F-плазмиды, на которой лучше всего и изучено. Гены, которые отвечают за поверхностное исключение, обозначают Sfx. Такие гены были идентифицированы в F-плазмиде и нескольких F-подобных плазмидах, например, R100. Известно пять групп плазмид, различных по признаку поверхностного исключения. Представителями этих групп являются плазмиды F, ColB2-K98, R1, R100 и pED208 соответственно. Иногда ряд F-подобных плазмид выделяют в шестую группу по признаку Sfx. Однако, даже если плазмиды относятся к разным группам по признаку Sfx, нельзя безоговорочно говорить о поверхностном исключении и невозможности конъюгации. Показано, что в большинстве случаев поверхностная несовместимость связана с особенностями половых пилей.
Конъюгация у других бактерий
Описанный выше механизм конъюгации относится к грамотрицательным бактериям. Вместе с тем к конъюгации способны многие грамположительные бактерии, от Streptomyces до Enterococcus. Во многих таких случаях механизм передачи ДНК принципиально схож с таковым у грамотрицательных бактерий. Однако в то время, как у грамотрицательных бактерий для конъюгации необходимы как минимум 20 генов, у грамположительных бактерий в конъюгации задействовано существенно меньше генов, иногда всего лишь пять. Поэтому конъюгативные плазмиды грамположительных бактерий зачастую мельче таковых у грамотрицательных. Кроме того, в силу особенностей строения клеточной стенки (толстый слой пептидогликана) в конъюгации у грамположительных бактерий не задействованы пили. Система конъюгации достаточно изучена у грамположительной бактерии Enterococcus faecalis. Некоторые штаммы этого вида секретируют растворимые феромоны пептидной природы, которые стимулируют экспрессию генов tra на конъюгативных плазмидах соседних клеток (любопытно, что в этом случае феромоны выделяют реципиенты генетического материала, а не наоборот, как это происходит обычно). У клеток-доноров, содержащих конъюгативную плазмиду, на поверхности находятся рецепторы к феромонам, причём разным плазмидам соответствуют разные феромоны и, соответственно, разные рецепторы. После связывания с рецептором феромон доставляется в цитоплазму специальным белком, и в цитоплазме феромон взаимодействует с белком TraA. В свободном виде этот белок подавляет экспрессию белков tra, а при связывании с феромоном утрачивает ингибиторные свойства, и экспрессия генов tra активируется. В результате образуется сложная белковая агрегация, соединяющая клетку-донора и реципиента.
Конъюгативные транспозоны
У E. faecalis возможна конъюгация, опосредованная не плазмидами, а транспозоном — Tn916. Он содержит сайт oriT, похожий на таковой у плазмид. В начале конъюгации Tn916 вырезается из бактериальной хромосомы двумя ферментами, которые сам и кодирует, — Int и Xis, которые родственны ферментам, обеспечивающим вырезание бактериофага λ. Далее он принимает кольцевую форму и становится похож на конъюгативную плазмиду, так как содержит oriT и гены tra. При этом даже в кольцевой форме Tn916 не имеет ориджина репликации, а потому неспособен к удвоению. Механизм переноса ДНК в случае Tn916 аналогичен таковому для конъюгативных плазмид. Когда в клетке-реципиенте образуется двуцепочечная кольцевая форма транспозона, фермент интеграза вставляет его в случайный участок бактериального генома. Tn916 является прототипом группы родственных конъюгативных транспозонов у грамположительных кокков, хотя конъюгативные транспозоны описаны и у грамотрицательных бактерий. Наряду с плазмидами, конъюгативные транспозоны тоже могут вносить свой вклад в распространение устойчивости к антибиотикам: так, Tn916 содержит ген устойчивости к тетрациклину.
Конъюгация и картирование бактериальной хромосомы
После открытия штаммов Hfr Элли Вольман и Франсуа Жакоб предложили новый метод картирования бактериальной хромосомы — метод прерывания скрещивания. Они смешивали в одной культуре F+ и F- клетки, несущие определённые хромосомные маркеры, инкубировали их некоторое время, потом путём резкого встряхивания, разрушающего контакты между клетками, прерывали конъюгацию и отбирали клетки, в которых произошла рекомбинация. Прерывая конъюгацию в разные временные промежутки и определяя, какие гены и на какой минуте после начала конъюгации передавались, можно установить порядок расположения генов, то есть картировать их на бактериальной хромосоме.
Литература
- Гигани О. Б. Плазмиды. — М.: РУСАЙНС, 2017. — 154 с. — ISBN 978-5-4365-1976-0.
- Инге-Вечтомов С. Г. Генетика с основами селекции. — СПб.: Издательство Н-Л, 2010. — 718 с. — ISBN 978-5-94869-105-3.
- Jeremy W. Dale, Simon F. Park. Molecular Genetics of Bacteria. — 4th Edition. — John Wiley & Sons, Ltd, 2004. — ISBN 0-470-85084-1.
- Joanne M. Willey, Linda M. Sherwood, Christopher J. Woolverton. Prescott's Principles of Microbiology. — 1st edition. — McGraw-Hill Higher Education, 2009. — P. 105—106. — 968 p. — ISBN 978-0-07-337523-6.