
| НАДН-убихинон-оксидоредуктаза | |
|---|---|
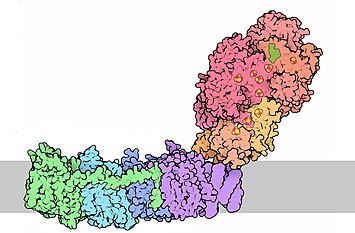 Серым цветом показана внутренняя мембрана митохондрии. Сверху — митохондриальный матрикс, снизу — межмембранное пространство. | |
| Идентификаторы | |
| Шифр КФ | 7.1.1.2 |
| Базы ферментов | |
| IntEnz | IntEnz view |
| BRENDA | BRENDA entry |
| ExPASy | NiceZyme view |
| MetaCyc | metabolic pathway |
| KEGG | KEGG entry |
| PRIAM | profile |
| PDB structures | RCSB PDB PDBe PDBj PDBsum |
| Gene Ontology | AmiGO • EGO |
| Поиск | |
| PMC | статьи |
| PubMed | статьи |
| NCBI | NCBI proteins |
НАДН-дегидрогена́зный ко́мплекс, также называемый ко́мплекс I или НАДН-убихино́н-оксидоредукта́за — первый мультибелковый комплекс дыхательной цепи переноса электронов. Множество копий комплекса расположены в мембранах прокариотических организмов, способных к кислородному дыханию и внутренних мембранах митохондрий эукариотических клеток. По отношению к белкам человека комплекс I часто называют НАДН-дегидрогеназой.
Этот комплекс играет центральную роль в процессах клеточного дыхания и окислительного фосфорилирования: почти 40 % протонного градиента для синтеза АТФ создаются именно этим комплексом. Комплекс I окисляет НАДН и восстанавливает одну молекулу убихинона, которая высвобождается в мембрану. На каждую окисленную молекулу НАДН комплекс переносит через мембрану четыре протона.
Комплекс І (НАДН-дегидрогеназа) был выделен из ряда объектов: митохондрий сердца быка, сахарной свеклы (Beta vulgaris), картофеля (Solanum tuberosum), бобов (Vicia faba), арабидопсиса (Arabidopsis thaliana) и риса (Oryza sativa), а также из митохондрий гриба нейроспоры Neurospora crassa и мембран кишечной палочки (Esherichia сoli).
Структурная организация комплекса I

У прокариот комплекс І состоит из 14 основных субъединиц, образующих ядро комплекса, без которого он не функционирует. Семь субъединиц чрезвычайно гидрофобны и локализованы в мембране, а семь относительно гидрофильных расположены вне мембраны. У эукариот в результате эволюции комплекс покрылся «шубой» из приблизительно 30 вспомогательных субъединиц, их количество может варьировать в зависимости от объекта. Так, у млекопитающих этот фермент состоит из 44 субъединиц, а у гриба Yarrowia lipolytica — из 48. В результате этой надстройки молекулярная масса комплекса I увеличилась почти вдвое: с ~550 кДа у бактерий до ~1 МДа в митохондриях.
Электронная микроскопия показала, что комплекс I (как из бактерий, так и из митохондрий) имеет характерную L-образную форму. Из-за этой формы, а также необычной, как бы мятой молекулярной поверхности, комплекс I получил от учёных прозвище «старый башмак». Гидрофобная «подошва» представлена белками, встроенными в мембрану, а гидрофильная часть — «лодыжка» — обращена в матрикс.
Четыре субъединицы убихинон-связывающего модуля совместно с субъединицами мембранной части фермента образуют сайт связывания убихинона, в котором он взаимодействует с железосерным кластером N2, принимает два электрона и восстанавливается до убихинола. Кластер N2, последний в ряду кластеров, по которым электроны от НАДН передаются к убихинону, приподнят над мембраной на ~15 Å. Сама полость, в которой связывается убихинон, имеет длину 30 Å и может вместить всю молекулу вместе с длинным гидрофобным хвостом из семи изопреновых единиц. Полость имеет узкий вход, так что длинная гидрофобная цепь вынуждена принять определённую конформацию, которая сохраняется на протяжении всей ферментативной реакции. Наличие такого длинного и узкого сайта связывания является характерной чертой комплекса I. Внутри полости убихинон взаимодействует с консервативными остатками тирозина и гистидина.
У грибов, животных и сосудистых растений по крайней мере семь из 44 субъединиц, которые образуют мембранный домен, кодируются митохондриальным геномом. У млекопитающих этих субъединиц ровно семь. У растений ДНК митохондрий кодирует девять субъединиц: кроме семи субъединиц, которые входят в состав гидрофобной части комплекса, в ней закодированы две субъединицы, гомологичные субъединицам 49 кДа и 30 кДа млекопитающих, а остальные компоненты находятся под контролем ядерных генов. Однако эти данные, полученные при исследовании пластомов картофеля и арабидопсиса, могут оказаться неверны для других видов растений, а количество закодированных в митохондриях субъединиц может изменяться от вида к виду. Так, у печёночника Marchantia polymorpha субъединица NAD7, гомологичная полипептиду 49 кДа, кодируется ядерным геномом и транспортируется в митохондрии, а соответствующий ген митохондриальной ДНК превратился в псевдоген и не функционален.
Исследования показали, что по своим свойствам комплекс I существенно не отличается у объектов животного и растительного происхождения. Однако у растений имеются некоторые специфические субъединицы, которые в некоторых случаях приводят к функциональным особенностям. Анализ комплекса I у Arabidopsis показывает, что более 30 % субъединиц специфичны только для растений. Например, частью мембранного модуля растительного комплекса I является так называемый γ-карбоангидразный структурный модуль и L-галактоно-1,4-лактондегидрогеназа, которая в то же время представляет собой последний фермент митохондриального пути биосинтеза аскорбиновой кислоты.
Таблица основных (коровых) субъединиц
| Коровые субъединицы | ||||||
|---|---|---|---|---|---|---|
| Корова | Человек | Кишечная палочка | Арабидопсис | Описание | Кофакторы | |
| Периферическая рука | ||||||
| Дегидрогеназный модуль | 75 кДа | NDUFS1 | NuoG | 75 кДа | транспорт электронов | Fe2S2 (N1b), 2 x Fe4S4 (N4 и N5), (N7) |
| 51 kDa | NDUFV1 | NuoF | 51 кДа | связывание НАДН, транспорт электронов | флавин, Fe4S4 (N3) | |
| 24 kDa | NDUFV2 | NuoE | 24 кДа | Несёт вспомогательный кластер N1a | Fe2S2 (N1a) | |
| Убихинон-связывающий модуль | 49 кДа | NDUFS2 | NuoD (NuoCD) | NAD7 | ||
| 30 kDa | NDUFS3 | NuoC | NAD9 | |||
| TYKY | NDUFS8 | NuoI | 28,5 кДа | транспорт электронов | 2 x Fe4S4 (N6a и N6b) | |
| PSST | NDUFS7 | NuoB | 20 кДа | транспорт электронов | Fe4S4 (N2) | |
| Мембранная рука | ||||||
| - | ND1 | MT-ND1 | NuoH | NAD1 | ||
| Протон-транслоцирующий модуль | ND2 | MT-ND2 | NuoN | NAD2 | антипортер-подобный белок | |
| ND3 | MT-ND3 | NuoA | NAD3 | |||
| ND4 | MT-ND4 | NuoM | NAD4 | антипортер-подобный белок | ||
| ND4L | MT-ND4L | NuoK | NAD4L | |||
| ND5 | MT-ND5 | NuoL | NAD5 | антипортер-подобный белок | ||
| ND6 | MT-ND6 | NuoJ | NAD6 | |||
Таблица вспомогательных субъединиц
Все митохондриальные комплексы I имеют множество дополнительных субъединиц, которые не являются необходимыми для каталитической активности и отличаются у разных видов. Очевидно, что они несут определённую функциональную нагрузку, поскольку мутации в них приводят к врождённым заболеваниям. Для некоторых субъединиц показано наличие определённых функций, так B16.6 (GRIM-19) участвует в апоптозе, а субъединица 39 кДа (NDUFA9) — в регуляции активности комплекса. Что касается остальных субъединиц, то сейчас активно обсуждается их возможная роль в регуляции, сборке, стабилизации и защите от активных форм кислорода. Следует упомянуть, что дополнительные субъединицы значительно увеличивают энергетические затраты клетки на синтез, сборку и деградацию комплекса. Такие затраты, тем не менее, могут окупится в случае эукариотической клетки, в которой процесс синтеза белка хорошо контролируется и в этом смысле доведён до совершенства. С другой стороны, если вспомогательные субъединицы необходимы для стабилизации комплекса I, остаётся непонятным каким образом бактериальные комплексы, которые состоят из минимально необходимого количества полипептидов, успешно функционируют без них. На данный момент у учёных нет чёткого ответа на эти вопросы.
| Вспомогательные субъединицы | ||||
|---|---|---|---|---|
| No. | Корова | Белок человека | Описание | Семейство белков Pfam |
| Субъединицы | ||||
| 15 | 13A | NDUFS6 | участвует в сборке и стабилизирует комплекс | Pfam PF10276 |
| 16 | B17.2 | NDUFA12 | Pfam PF05071 | |
| 17 | AQDQ | NDUFS4 | возможно фосфорилируется ПКА, стабилизирует комплекс во время сборки |
Pfam PF04800 |
| 18 | 39kDa | NDUFA9 | соединяет мембранный и периферический домены, участвует в активации/деактивации комплекса |
Pfam PF01370 |
| 19 | ACPM | ACPM_HUMAN | ацил-переносящий белок | Pfam PF00550 |
| 20 | B8 | NDUA2_HUMAN | Pfam PF05047 | |
| 21 | MFWE | NDUFA1 | цАМФ зависимое фосфорилирование ПКА | Pfam PF15879 |
| 22 | B12 | NDUFB3 | Pfam PF08122 | |
| 23 | AB13 | NDUFA5 | участвует в активации/деактивации комплекса | Pfam PF04716 |
| 24 | B14 | NDUFA6 | Pfam PF05347 | |
| 25 | B14.7 | NDUFA11 | Pfam PF02466 | |
| 26 | ESSS | NDUFB11 | цАМФ зависимое фосфорилирование ПКА | Pfam PF10183 |
| 27 | PFFD | NDUFS5 | Pfam PF10200 | |
| 28 | B15 | NDUFB4 | Pfam PF07225 | |
| 29 | B16.6 | NDUFA13 | участвует в апоптозе | Pfam PF06212 |
| 30 | B18 | NDUFB7 | Pfam PF05676 | |
| 31 | PGIV | NDUFA8 | Pfam PF06747 | |
| 32 | B22 | NDUFB9 | Pfam PF05347 | |
| 33 | PDSW | NDUFB10 | фосфорилируется Src в раковых клетках | Pfam PF10249 |
| 34 | ASHI | NDUFB8 | Pfam PF05821 | |
| 35 | B14.5B | NDUFC2 | в норме фосфорилирована | Pfam PF06374 |
| 36 | AGGG | NDUFB2 | Pfam PF14813 | |
| 37 | B14.5A | NDUFA7 | в норме фосфорилирована | Pfam PF07347 |
| 38 | B9 | NDUFA3 | Pfam PF14987 | |
| 39 | MLRQ | NDUA4_HUMAN | Pfam PF06522 | |
| 40 | SGDH | NDUFB5 | Pfam PF09781 | |
| 41 | MNLL | NDUFB1 | Pfam PF08040 | |
| 42 | KFYI | NDUFC1 | в норме фосфорилирована | Pfam PF15088 |
| 43 | 42kD | NDUFA10 | Pfam PF01712 | |
| 44 | NDUFA4L2 | NDUFA4L2 | Pfam PF15880 | |
| 45 | NDUFV3 | NDUFV3 | - | |
| 46 | NDUFB6 | NDUFB6 | Pfam PF09782 | |
|
Вспомогательные белки, осуществляющие сборку | ||||
| 48 | NDUFAF2 | MIMIT_HUMAN | фактор сборки комплекса 2 | Pfam PF05071 |
| 49 | NDUFAF3 | NDUF3_HUMAN | фактор сборки комплекса 3 | Pfam PF05071 |
| 50 | NDUFAF4 | NDUF4_HUMAN | фактор сборки комплекса 4 | Pfam PF06784 |
Некоторые дополнительные субъединицы фосфорилируются различными киназами, чего никогда не происходит с коровыми субъединицами. Предположителагается, что таким образом происходит регуляция работы комплекса. Как одна из субъединиц в комплексе присутствует ацил-переносящий белок (NDUFAB1) с фосфорилированной пантотеновой кислотой в качестве в простетической группы. Предполагают, что он участвует в синтезе липоевой кислоты, репарации повреждённых липидов мембраны или модифицирует другие белки остатками миристиновой кислоты. Следует отметить, что функционирование этого белка не зависит от прямого физического контакта с комплексом I и значительная его часть присутствует в свободном виде внутри митохондриального матрикса.
Кофакторы

Все простетические группы НАДН-дегидрогеназного комплекса (один флавинмононуклеотид и от 8 до 9 железосерных кластеров) находятся в периферическом водорастворимом домене. У млекопитающих, как и у всех позвоночных, их восемь. Семь кластеров образуют электрон-транспортную цепь длинной в ~96 Å от ФМН до места связывания убихинона. На основе современных данных считается, что перенос электрона происходит по следующему пути: НАДН → ФМН → N3 → N1b → N4 → N5 → N6a → N6b → N2 → Q. Сначала происходит передача двух электронов на флавин, а затем они по одному передаются через цепочку кластеров к сайту связывания хинона и восстанавливают его до состояния Q−2. Кластер N1a располагается недалеко от флавинового кофактора и на некотором расстоянии от магистральной цепи переноса электрона. Этот кластер высококонсервативен у разных видов; полагают, что он осуществляет контроль скорости транспорта электрона внутри комплекса, перебрасываясь электроном с ФМН. Существует модель, согласно которой один из электронов с флавина идёт по магистральному пути на хинон, а другой запасается в кластере N1a и позже возвращается в основную цепь, через флавосемихинон. Возможно, такой механизм позволяет снизить образование активных форм кислорода на восстановленном флавине. К тому же, это позволяет стабилизировать (до миллисекунды) состояние, когда последний кластер N2 восстановлен, но нет второго электрона, чтобы завершить восстановление убихинона. Такое состояние может быть необходимым для конформационных изменений, сопряжённых с транспортом протонов.
Часть кластеров в цепи (N3, N4 и N6a) имеют высокий окислительно-восстановительный потенциал (редокс-потенциал) на уровне —0,25 В, в то время как три других (N1b, N5 и N6b) имеют более низкие потенциалы. В результате редокс-потенциал на пути электрона меняется наподобие американских горок. Такая кривая изменения энергетического состояния характерна для многих окислительно-восстановительных ферментов: она позволяет оптимизировать скорость транспорта электрона и добиться эффективного переноса энергии.
Кластер N5 имеет очень низкий потенциал и лимитирует скорость общего потока электронов по всей цепи. Вместо обычных для железосерных центров лигандов (четырёх остатков цистеина) он скоординирован тремя остатками цистеина и одним остатком гистидина, а также окружён заряженными полярными остатками, хотя и находится в глубине фермента.
Необычные лиганды имеет и терминальный кластер цепи — N2. Его редокс-потенциал самый высокий из всех кластеров (от —0,1 до —0,15 В). Он связан с четырьмя последовательно расположенными в полипептидной цепи остатками цистеина, что создаёт напряжённую конформацию. Из-за этого при его восстановлении происходят конформационные изменения соседних цепей, возможно, связанные с транспортом протона.
Кластер N7 присутствует только в комплексе I некоторых бактерий. Он значительно удалён от остальных кластеров и не может обмениваться с ними электронами, так что по-видимому, является реликтом. В некоторых бактериальных комплексах, родственных комплексу I, между N7 и остальными кластерами обнаружены четыре консервативных остатка цистеина, а комплексе I бактерии Aquifex aeolicus был обнаружен дополнительный Fe4S4 кластер, соединяющий N7 с остальными кластерами. Из этого следует вывод, что у A. aeolicus комплекс I, кроме НАДН, может использовать иной донор электронов, который передаёт их через N7.
Сборка митохондриального комплекса I
Митохондриальный комплекс I образует с дыхательными комплексами III и IV суперкомплексы, именуемые респирасомами. В митохондриях млекопитающих и человека около 90 % комплекса находится в составе респирасом. Также на митохондриях из молодых корневищ бамбука показано, что 90 % от общего количества комплекса I собрано в респирасомы, а у Arabidopsis — в суперкомплекс I—III2. Накоплено достаточно доказательств того, что присутствие респирасом необходимо для стабильности и функционирования комплекса I, который в отсутствии комплексов III или IV нестабилен. Например, на мутантных клетках человека показано, что комплекс I необходим для формирования комплекса III, и с другой стороны, утрата комплекса III приводит к потере комплекса I. Кроме того, в ряде исследований на животных клетках приводятся доказательства того, что для стабильности комплекса I необходимы комплексы IV и димер комплекса III.
Недавно на культуре клеток человека было показано, что комплексы IV и III необходимы для сборки полноценного комплекса I, при этом сам не до конца собранный комплекс служит основой для образования респирасом. Наличие в респирасоме комплексов IV и III нужно для присоединения к комплексу I каталитических субъединиц НАДН-дегидрогеназного модуля, которые полностью активируют комплекс и всю респирасому.
Реакция
НАДН-дегидрогеназный комплекс окисляет НАДН, образовавшийся в матриксе в ходе цикла трикарбоновых кислот. Электроны от НАДН используются для восстановления мембранного переносчика, убихинона Q, который переносит их к следующему комплексу электрон-транспортной цепи митохондрий, комплексу III или цитохром-bc1-комплексу.
НАДН-дегидрогеназный комплекс работает как протонная помпа: на каждый окисленный НАДН и восстановленный Q через мембрану в межмембранное пространство перекачиваются четыре протона:
- НАДН + H+ + Q + 4H+in → НАД+ + QH2 + 4H+out
Образовавшийся в ходе реакции электрохимический потенциал используется для синтеза АТФ. Любопытно, что реакция, катализируемая комплексом I, обратима, этот процесс называется аэробное сукцинат-индуцированное восстановление НАД+. В условиях большого потенциала на мембране и избытка восстановленных убихинолов комплекс может восстанавливать НАД+ с использованием их электронов и пропускать протоны обратно в матрикс. Этот феномен обычно наблюдается, когда много сукцината, но мало оксалоацетата или малата. Восстановление убихинона осуществляется ферментами сукцинатдегидрогеназой, глицерол-3-фосфатдегидрогеназой или митохондриальной дигидрооротатдегидрогеназой. В условиях высокого протонного градиента сродство комплекса к убихинолу повышается, а редокс-потенциал убихинола снижается благодаря росту его концентрации, что и делает возможным обратный транспорт электронов по электрическому потенциалу внутренней мембраны митохондрий к НАД. Данный феномен удалось наблюдать в лабораторных условиях, но неизвестно, имеет ли он место в живой клетке.
Механизм транспорта протонов

На начальных этапах исследования комплекса I широко обсуждалась модель, основанная на предположении, что в комплексе оперирует система, похожая на Q-цикл. Однако позднейшие исследования не обнаружили в комплексе I каких-либо внутренне связанных хинонов и полностью опровергли эту гипотезу.

НАДН-дегидрогеназный комплекс, по-видимому, имеет уникальный механизм транспорта протонов посредством конформационных изменений самого фермента. Субъединицы ND2, ND4 и ND5 называются антипорт-подобными, поскольку они гомологичны друг другу и бактериальным Mrp Na+/H+ антипортам. Эти три субъединицы образуют три основных протонных канала, которые состоят из консервативных остатков заряженных аминокислот (в основном лизина и глутамата). Четвёртый протонный канал образован частью субъединицы Nqo8 и малыми субъединицами ND6, ND4L и ND3. Канал сходен по строению с аналогичными каналами антипорт-подобных субъединиц, но содержит необычно много плотно упакованных остатков глутамата со стороны матрикса, за что и получил название E-канал (латинское E используется как стандартное обозначение глутамата). От С-конца субъединицы ND5 отходит удлинение, состоящее из двух трансмембранных α-спиралей, соединённых необычно протяжённой (110 Å) α-спиралью (HL), которая, проходя по стороне комплекса, обращённой в матрикс, физически соединяет все три антипорт-подобные субъединицы, и возможно, участвует в сопряжении транспорта электронов с конформационной перестройкой. Ещё один сопрягающий элемент, βH, образован серией перекрывающихся β-шпилек и α-спиралей, он расположен на противоположной, периплазматической стороне комплекса.
До сих пор окончательно неизвестно, как именно транспорт электронов сопряжён с переносом протонов. Полагают, что мощный отрицательный заряд кластера N2 может расталкивать окружающие полипептиды, вызывая тем конформационные изменения, которые неким образом распространяются на все антипорт-подобные субъединицы, расположенные довольно далеко друг от друга. Другая гипотеза предполагает, что изменение конформации вызывает в необычно длинном сайте связывания убихинона стабилизированный убихинол Q−2 с крайне низким редокс-потенциалом и отрицательным зарядом. Неизвестными остаются и многие детали кинетики конформационных изменений и сопряжённого с ними транспорта протонов.
Активная и неактивная формы
Эукариотический НАДН-дегидрогеназный комплекс существует в двух чётко различимых формах: одна полностью функционирующая, так называемая активная или A-форма, и вторая, каталитически неактивная или D-форма. Если фермент находится при повышенных, но всё ещё физиологических температурах (> 30 °C) в отсутствие субстрата, фермент переходит в D-форму. Она каталитически неактивна, но может активироваться субстратом (НАДН и убихиноном, на который можно сбрасывать электроны). После одного или нескольких ферментативных циклов комплекс становится активным, а скорость реакции возрастает. Такой переход обнаружен только у позвоночных и грибов, но не у беспозвоночных или бактерий. Растительные комплексы не исследовались. В присутствии бивалентных катионов (Mg2+, Ca2+) или в щелочном pH активация происходит значительно дольше, а свободная пальмитиновая кислота сильно увеличивает частоту перехода из активной в деактивированную форму.
 |
 |
|||||||||
|
Комплекс I из быка и Yarrowia lipolytica
| ||||||||||
Высокая энергия активации (270 кДж/моль) перехода из A- в D-форму указывает, что в комплексе происходит значительная конформационная перестройка. До настоящего момента единственным выявленным различием между двумя формами остаётся количество остатков цистеина на поверхности фермента. Согласно последним данным, в этом процессе участвуют субъединицы, расположенные рядом с хинон-связывающим сайтом: 39 кДа, ND3 и ND1. Обработка D-форм комплекса I специальными реагентами (N-этилмалеимидом или реактивом Эллмана) необратимо блокирует эти важные остатки цистеина, делая невозможной повторную активацию фермента. Любопытно, что A-форма комплекса I нечувствительна к сульфгидрилам, что указывает на то, что остатки цистеина спрятаны глубоко в белке. В свою очередь деактивированная форма восприимчива к ингибированию нитрозотиолами и пероксинитритом.
Конформационные изменения комплекса I имеют большое физиологическое значение. После гипоксии восстановление уровня кислорода может привести к всплеску окисления НАД(Ф)Н и генерации активных форм кислорода, которые могут повредить митохондрии и вызвать некроз тканей. Переход из активной в неактивную форму комплекса происходит при патологических состояниях, когда число оборотов фермента снижено при нормальной, физиологической температуре тела, например при гипоксии, ишемии или повышении соотношения оксид азота (NO)/кислород в тканях (так называемая метаболическая гипоксия). Таким образом комплекс I предотвращает окисление остальных дыхательных комплексов, когда происходит восстановление уровня кислорода. Кроме того, неактивная форма не способна к обратному транспорту электронов, что снижает образование АФК.
Эволюционное происхождение


НАДН-дегидрогеназный комплекс относится к семейству мембранных оксидоредуктаз из класса NiFe-гидрогеназ, которые у анаэробных бактерий и архей сопрягают реакцию окисления субстрата и восстановления водорода с транспортом протонов. На основе данных о гомологии белков можно сделать вывод, что комплекс возник в результате объединения двух ранее существовавших комплексов из разных, неродственных белковых семейств. НАДН-дегидрогеназный и убихинон-связывающий модули произошли из растворимой NiFe-гидрогеназы, которая окисляла НАДН и восстанавливала водород, а гидрофобная мембранная «подошва» комплекса, перекачивающая протоны, возникла из Na+/H+-антипортов Mrp.
Слияние растворимой гидрогеназы и белков-антипортов привело к возникновению большого количества мембранных гидрогеназ и дегидрогеназ, которые могли позднее эволюционировать в комплекс I. Трёхмерная структура этих ферментов, вероятно, схожа с таковой у комплекса I. К дегидрогеназам можно отнести архейный комплекс Fpo из 11 субъединиц, который окисляет кофактор F420, связанный с водородом, и восстанавливает метанофеназин (аналог убихинона), перекачивая через мембрану один протон на два электрона. У этого фермента нет НАДН-дегидрогеназного модуля. К группе гидрогеназ относятся формиатводородлиазы из Escherichia coli: формиатводородлиаза-1 из семи субъединиц и формиатводородлиаза-2 из десяти. Оба фермента окисляют формиат, восстанавливая водород с переносом нескольких протонов через мембрану.
Самый простой из белков-родственников комплекса I — гидрогеназа Ech (англ. E. coli hydrogenase-3-type hydrogenase) археи Methanosarcina barkeri. Она состоит из всего шести субъединиц и перекачивает один протон в результате окисления ферредоксина с восстановлением молекулы водорода. Ech содержит минимальный набор субъединиц (гомологичных комплексу I), необходимый для сопряжения реакции окисления с транспортом протонов.
Кроме того, комплекс I обнаружен в хлоропластах в виде НАДН-дегидрогеназного комплекса хлоропластов. Его точное строение и функции пока неизвестны.
Образование активных форм кислорода
Комплекс I в процессе своей работы образует активные формы кислорода. Обычно это супероксид (а также пероксид водорода), и образуется он по крайней мере двумя путями. В ходе прямого транспорта электронов, в процессе дыхания, образуется очень малое количество супероксида (вероятно, менее 0,1 % от всего потока электронов переносятся на кислород).
Во время обратного транспорта электронов, который происходит в условиях аэробного сукцинат-индуцированного восстановление НАД+, комплекс I, возможно, становится самым активным местом образования супероксида: до 5 % электронов идут на восстановление кислорода.
Супероскид образуется в НАДН-дегидрогеназный комплексе в результате переноса одного электрона с ФМНН2 к O2. Образовавшийся в результате радикал флавина нестабилен и переносит оставшийся электрон на железосерные кластеры. Уровень образования супероксида определяется соотношением НАДН/НАД+; в условиях когда восстановлено небольшое количество НАД, НАД+ успешно конкурирует за электроны с кислородом.
Ингибиторы
Наиболее изученный ингибитор комплекса I — ротенон (широко применяемый как органический пестицид). Ротенон и ротеноиды — это изофлавоноиды, которые присутствуют в корнях нескольких родов тропических растениях таких как Антония (Loganiaceae), Derris и Lonchocarpus (Fabaceae). Ротенон давно используется как инсектицид и рыбный яд, так как митохондрии насекомых и рыб особенно к нему чувствительны. Известно, что коренные жители Французской Гвианы и другие индейцы Южной Америки использовали ротенон-содержащие растения для рыболовства уже в XVII веке. Ротенон взаимодействует с сайтом связывания убихинона и конкурирует с основным субстратом. Было показано, что долгосрочное системное подавление комплекса I ротеноном может индуцировать селективное отмирание дофаминергических нейронов (секретирующих в качестве нейротрансмиттера дофамин). Схожим образом действует и пиерицидин А, ещё один мощный ингибитор комплекса I, структурно схожий с убихиноном. К этой же группе относится и амитал натрия — производное барбитуровой кислоты.
Несмотря на более чем 50-летнее изучение комплекса I, так и не удалось обнаружить ингибиторы, блокирующие перенос электронов внутри комплекса. Гидрофобные ингибиторы, такие как ротенон или пиерицидин, просто прерывают перенос электрона с терминального кластера N2 на убихинон.

Ещё одно вещество, блокирующее комплекс I — это аденозиндифосфатрибоза, конкурентный ингибитор в реакции окисления НАДН. Он связывается с ферментом в сайте связывания нуклеотида (ФАД).
К одним из самых сильных ингибиторов комплекса I относится семейство ацетогенинов. Показано, что эти вещества образуют химические сшивки с субъединицей ND2, что косвенно указывает на роль ND2 в связывании убихинона. Любопытно отметить, что ацетогенин роллиниастатин-2 стал первым из обнаруженных ингибиторов комплекса I, который связывается в другом месте, нежели ротенон.
Умеренным ингибиторным эффектом обладает антидиабетический препарат метформин; по-видимому, данное свойство препарата лежит в основе механизма его действия.
Патологии
Мутации в генах субъединиц комплекса I могут привести к митохондриальным заболеваниям, например, к синдрому Лея. Точечные мутации митохондриальных субъединиц этого комплекса также могут стать причиной оптической нейропатии Лебера. Есть доказательства, что дефекты в структуре комплекса I могут играть роль в этиологии болезни Паркинсона, возможно, из-за образования активных форм кислорода. Так, было показано, что у культур клеток больных болезнью Паркинсона увеличена утечка протонов в комплексе I, что снижает максимальную дыхательную ёмкость. У растений мутации по комплексу І описаны у табака (Nicotiana silvestris) и кукурузы (Zea mays): мутации сопровождались патологией пыльцы и приводили к цитоплазматической мужской стерильности.
Недавние исследования выявили необычную роль комплекса I в работе мозга. Активность этого фермента значительно снижена у пациентов с биполярным расстройством, но остаётся в норме у пациентов с депрессией или шизофренией. У пациентов с биполярным расстройством в префронтальной коре наблюдалось повышенное окисление и нитрирование белков. Эти результаты делают комплекс I мишенью для будущих терапевтических исследований биполярного расстройства.
Воздействие пестицидов, блокирующих комплекс I, может приводить к далеко идущим последствиям. Например, продолжительное воздействие низких концентраций органофосфата и пестицида дихлофоса вызывает дисфункцию печени. Дихлофос изменяет активность комплексов I и II, что приводит к замедлению транспорта электронов и снижению синтеза АТФ.
Роль комплекса I в процессах старения
Данные многочисленных исследований свидетельствуют о том, что митохондрии, и в частности комплексы I и II играют ключевую роль в процессах, влияющих на старение и на продолжительность жизни. Предполагается что замедление при старении синтеза и восполнения белков приводит к сбою в стехиометрии дыхательных субъединиц. Это в свою очередь вызывает нарушение эффективности функционирования комплекса I и повышение митохондриального окислительного стресса, что наиболее ярко выражено в мышечной ткани.
Вставка в дополнение к комплексу I в геном дрозофилы альтернативной НАДН-дегидрогеназы Ndi1 дрожжей, состоящей всего из одной субъединицы, приводило к восстановлению нормального уровня окисления внутримитохондриальной НАДН и значительному продлению жизни этой мухи независимо от ограничения калорийности её диеты.
См. также
Источники
Литература
- Физиология растений / Под ред. И. П. Ермакова. — М.: Академия, 2005. — 634 с.