

Презентация антиге́на (англ. antigen presentation) — процесс предъявления T-лимфоциту фрагмента антигена с целью запуска T-клеточного ответа. Поскольку T-клетки не распознают нативные антигены, то нативные антигены предварительно фрагментируются антигенпрезентирующей клеткой, и фрагменты выставляются на её поверхности в связанном с главным комплексом гистосовместимости (MHC) виде, чтобы они могли быть распознаны T-клеточными рецепторами. В случае вирусной или бактериальной инфекции антигенпрезентирующая клетка выставляет на своей поверхности эндогенные или экзогенные пептидные фрагменты, полученные от исходного антигена, в виде комплекса с MHC. Существуют два класса молекул MHC, которые различаются происхождением фрагментов антигена, которые с ними связываются: молекулы MHC I класса (MHC-I) связывают пептидные фрагменты, происходящие из цитозоля клетки (то есть эндогенные), а молекулы MHC II класса (MHC-II) связывают фрагменты экзогенного происхождения, которые появились в результате эндоцитоза исходного антигена и его последующего расщепления. Каждая T-клетка способна распознавать от нескольких десятков до нескольких сотен фрагментов одного и того же антигена, хотя на поверхности антигенпрезентирующей клетки могут быть экспонированы тысячи других пептидных фрагментов, поскольку одна и та же молекула MHC способна связывать самые разнообразные пептиды.
В отличие от T-клеток, B-клетки способны с помощью B-клеточных рецепторов связывать интактные антигены с их нативной структурой, а не их линейные пептидные фрагменты.
Презентация внутриклеточных антигенов
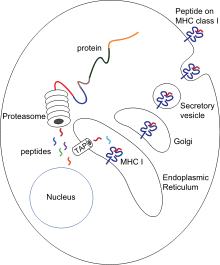
Цитотоксические T-клетки, также известные как T-киллеры, отличаются экспрессией корецептора CD8 и специализируются на том, что вызывают программируемую клеточную гибель других клеток. Когда T-киллеры обнаруживают признаки болезни — вирусной инфекции, инфекции, вызванной внутриклеточными бактериями, или злокачественные клетки — они запускают гибель потенциально опасных клеток (то есть клеток, заражённых вирусом или внутриклеточными бактериями, а также злокачественных клеток). Все клетки, имеющие ядра, а также тромбоциты экспрессируют MHC I класса, молекулы которого располагаются в клеточной мембране. Антигены, подвергшиеся процессингу в этих клетках, выставляются на их поверхности в комплексе с молекулами MHC I класса.
В ходе процессинга антигена исходный антиген расщепляется на короткие пептиды цитозольными протеазами. Далее полученные пептидные фрагменты доставляются в эндоплазматический ретикулум (ЭПР) при участии белков теплового шока и транспортёром, связанным с процессингом антигена (англ. transporter associated with antigen processing, TAP). TAP переносит цитозольные пептиды в люмен ЭПР с затратой АТФ. В люмене ЭПР имеется несколько шаперонов, связанных со сборкой MHC-I, а именно, кальнексин, кальретикулин и тапазин. Пептиды загружаются на MHC-I через пептид-связывающую бороздку, которая расположена между двумя α-спиралями в нижней части доменов α1 и α2 MHC-I. Когда от комплекса MHC-I с фрагментом антигена (pMHC-I) отделяется тапазин, pMHC-I выходит из ЭПР и транспортируется к поверхности клетки внутри экзоцитозных везикул.
Наивные CD8+ T-клетки не могут непосредственно убить инфицированную или злокачественную клетку. Они должны быть активированы комплексом pMHC-I, который несёт антигенпрезентирующая клетка. Антиген может быть презентирован непосредственно по механизму, описанному выше, или с помощью кросс-презентации от инфицированных или неинфицированных клеток. После взаимодействия pMHC-I с T-клеточным рецептором на поверхности наивной CD8+ T-клетки при участии костимулирующих сигналов и/или цитокинов T-клетка активируется и мигрирует в периферические ткани, где убивает клетки, экспрессирующие тот антиген, которые ей был представлен.
В ходе кросс-презентации молекулы MHC-I презентируют внеклеточные антигены, которые обычно презентируются MHC-II. Такой способностью обладают MHC-I некоторых антигенпрезентирующих клеток, в особенности, плазмацитоидные дендритные клетки. Кросс-презентация происходит тогда, когда сама антигенпрезентирующая клетка не инфицирована сама, но запускает локальный противовирусный или противоопухолевый иммунный ответ без миграции в близлежащие лимфоузлы.
Презентация внеклеточных антигенов
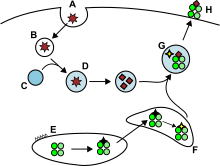
Антигены из внеклеточного пространства, а также некоторые эндогенные антигены заключаются в эндоцитозные везикулы, доставляются на поверхность клетки и связываются молекулами MHC-II. Их распознают T-хелперы, экспрессирующие корецептор CD4. Только дендритные клетки, B-клетки и макрофаги экспрессируют MHC-II в достаточном количестве, поэтому экспрессия MHC-II более специфична, чем экспрессия MHC-I.
Антигенпрезентирующие клетки, как правило, поглощают антиген из внеклеточного пространства с помощью эндоцитоза, а также пиноцитоза, макроаутофагии, эндосомной микроаутофагии или аутофагии, опосредованной шаперонами. В случае эндоцитоза антигены попадают в везикулы, называемые эндосомами. В процессинге антигена задействованы три компартмента: ранние эндосомы, поздние эндосомы, или эндолизосомы, и лизосомы, в которых антигены гидролизуются лизосомными ферментами (кислыми гидролазами, глюкозидазами, протеазами, липазами). Расщеплению способствует постепенное понижение pH в везикулах. Главную роль в расщеплении антигена играют эндосомные гидролазы катепсины, разрезающие антиген на короткие пептиды.
Молекулы MHC-II транспортируются из ЭПР в погрузочный компартмент для MHC-II в месте с полипептидом, известным как инвариантная цепь (Ii, CD74). Специфическая молекула класса MHC-II (HLA-DO и HLA-DM) катализируют обмен части CD74, известный как CLIP-пептид, на фрагмент антигена. Образовавшийся комплекс пептид-MHC-II (pMHC-II) транспортируется в клеточную мембрану, где фрагмент антигена презентируется T-хелперам в лимфоузлах.
Мигрируя к лимфоидным тканям с помощью хемотаксических сигналов, антигенпрезентирующие клетки проходят процесс созревания, в ходе которого они утрачивают способность к фагоцитозу и становятся более способными к взаимодействию с T-клетками и презентации им антигена. Как и в случае T-киллеров, для активации T-хелперов, помимо pMHC-II, необходимы дополнительные костимулирующие сигналы.
Презентация нативных антигенов B-клеткам
B-клеточные рецепторы, располагающиеся на поверхности B-клеток, связываются с нерасщеплёнными антигенами, имеющими нативную конформацию, а не с линейными пептидными фрагментами антигенов в комплексе с MHC. Большие комплексы интактных антигенов презентируются B-клеткам в лимфоузлах фолликулярными дендритными клетками в форме иммунных комплексов. Некоторые антигенпрезентирующие клетки экспрессируют сниженное количество лизосомных ферментов, благодаря чему могут презентировать B-клеткам нерасщеплённые антигены.