
Эволюционная биология развития (англ. evolutionary developmental biology, evo-devo) — область биологии, которая, сравнивая онтогенез различных организмов, устанавливает родственные связи между ними и вскрывает развитие онтогенетических процессов в ходе эволюции. Она рассматривает происхождение и эволюцию эмбрионального развития; как изменения в онтогенетическом развитии приводят к возникновению новых признаков; роль фенотипической пластичности в эволюции; как экология влияет на развитие и эволюционные изменения; онтогенетическую основу гомоплазии и гомологии.
Хотя интерес к связи между онтогенезом и филогенезом возник ещё в XIX веке, современная биология развития получила новый импульс от открытий в области генетического регулирования эмбрионального развития в модельных организмах. Общие гипотезы всё ещё нуждаются в тщательной проверке, поскольку организмы очень различны по своему строению.
Несмотря на это, уже сейчас концепция «evo-devo» даёт понять, что подобно тому, как эволюция создаёт новые гены из частей старых, она же изменяет процессы индивидуального развития, создавая новые и даже новаторские структуры на основе старых генных сетей (подобно тому, как костная структура челюсти дала начало слуховым косточкам среднего уха) или сохраняя похожие программы развития во множестве различных организмов (например, генное регулирование развития глаза сходно у моллюсков, членистоногих и позвоночных). Изначально основной интерес был прикован к очевидной гомологичности клеточных и молекулярных механизмов, регулирующих план строения и развитие органов. Хотя более современный подход учитывает перестройки развития, связанные с видообразованием.
Основные принципы
Дарвиновская теория эволюции основана на трёх принципах: естественный отбор, наследственность и изменчивость. В то время, когда Дарвин создавал свою теорию, представления о наследственности и изменчивости были весьма туманны. Однако в 1940-х годах биологи использовали менделевские принципы генетики для объяснения эволюционного процесса, результатом чего стал «современный синтез». Но только в 1980-90-х годах, после того, как был накоплен и подробно изучен массив сравнительных данных о молекулярных последовательностях различных видов организмов, возникло понимание молекулярной основы механизмов индивидуального развития.
В настоящее время твёрдо установлено, как происходят мутации. Тем не менее, механизмы развития выяснены недостаточно, чтобы объяснить, какие виды фенотипической изменчивости могут возникать в каждом поколении из изменчивости на генетическом уровне. Эволюционная биология развития изучает, как динамика развития определяет фенотипическую изменчивость, возникающую на основе генетической изменчивости, а также, как она воздействует на эволюцию фенотипа. В то же время эволюционная биология развития изучает также эволюцию индивидуального развития.
Таким образом, истоки эволюционной биологии развития берут начало как в совершенствовании методов молекулярной биологии применительно к биологии развития, так и из признания ограниченности классического неодарвинизма применительно к эволюции фенотипа. Некоторые исследователи видят в концепции «evo-devo» расширение «современного синтеза» включением в него данных, полученных молекулярной генетикой и биологией развития. Другие, опираясь на обнаруженные расхождения между генотипом и фенотипом, а также на эпигенетические механизмы развития, бросают открытый вызов синтетической теории эволюции.
Эволюционная биология развития пока ещё не стала единой дисциплиной, но она существенно отличается от предшествующих направлений эволюционной теории своей сосредоточенностью на некоторых ключевых идеях. Одна из них — модульность в организации растений и животных, которые состоят из онтогенетически и анатомически отдельных частей. Часто такие части повторяются несколько раз в одном организме, как, например, пальцы, рёбра и сегменты тела. «Evo-devo» ищет генетические и эволюционные основы разделения эмбриона на отдельные модули, а также частично независимого развития таких модулей.
Другая центральная идея состоит в том, что некоторые продукты экспрессии генов функционируют как переключатели, тогда как другие действуют как переносчики сигнала. Гены кодируют структуру белков, некоторые из которых выполняют структурную функцию в клетках, а другие, такие как энзимы, регулируют различные пути биохимических превращений в организме. Большинство биологов, придерживающихся принципов синтетической теории, видят в организме непосредственное отражение его генетической составляющей. А изменение существующих или эволюция новых биохимических путей (и, в конечном счёте, возникновение новых видов организмов) основывается на определённых генетических мутациях. Но 1961 году Жак Моно, Жан-Пьер Шанже (англ. Jean-Pierre Changeux) и Франсуа Жакоб открыли у бактерий кишечной палочки ген, который функционирует, только будучи «включенным» средовым стимулом. Позже учёные открыли специфичные гены у животных, включая подгруппу генов, содержащих последовательность гомеобокса, так называемые "хокс-гены" (Hox-genes), которые действуют как переключатели других генов и могут быть индуцированы другими генетическими продуктами, морфогены, которые действуют аналогично внешним стимулам в случае бактерий. Эти открытия приковали внимание биологов к тому факту, что гены избирательно включаются и выключаются, а не пребывают в постоянной активности, и что совершенно разные организмы (например, дрозофила и человек) могут использовать похожие гены в эмбриогенезе.
Точно так же морфология организма может находиться под влиянием мутаций, происходящих в промоторных участках генов, в ДНК-последовательностях, с которыми продукты некоторых генов связываются и контролируют активность тех или иных генов. Это предполагает, что ключевое различие между разными видами (даже разными отрядами и типами) может быть обусловлено в меньшей мере составом их генетических продуктов, чем пространственными и временны́ми различиями экспрессии их постоянных генов. Подразумевается, что крупные эволюционные изменения в морфологии связаны скорее с изменениями в генной регуляции, чем с эволюцией новых генов, возможно, что Hox-гены и другие «переключатели» могут играть важную роль в эволюции, что противоречит синтетической теории эволюции.
Ещё одно направление «evo-devo» — пластичность онтогенеза, которая служит основой того факта, что фенотип организма не полностью определяется его генотипом. Если формирование фенотипов обусловлено и зависит от внешних или средовых влияний, эволюция может протекать по пути «первичного фенотипа» с генетическими изменениями, которые скорее следуют, чем инициируют, появление морфологических и других фенотипических новшеств. Аргументы в пользу этого приведены Мэри Джейн Вест-Эберхард (англ. Mary Jane West-Eberhard) в её книге 2003 года «Developmental plasticity and evolution».
Глубокая гомология

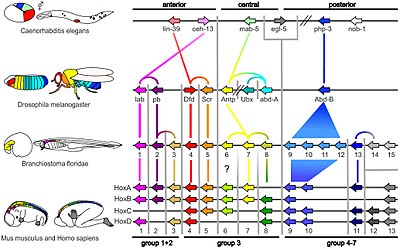
Глубокая гомология - это терминобозначающий процессы роста и развития и предварительной разметки организмов, при которых они регулируются генетическими механизмами, гомологичными и консервативными у широкого круга видов. Как пример это развитие организмов из зародышевых клеток, которые вплоть до зиготы могут стать любыми клетками в организме. Этот принцип заложен в развитии и китов и бабочек и медуз и слонов. Многие из этих организмов имеют одни и те же структурные гены для построения белков тела, так называемые hox гены или гомеозисные гены. Данные гены являются базовыми, одними из самых древних и общими для всех животных на планете.
У жирафов нет гена длинной шеи, у слонов нет гена большого тела. Их тела вырастают через систему переключения настроек в ДНК, эта система точно контролирует развитие тех или иных частей тела и черт внешности. При этом место где будет развиваться ткань, орган, кость так же отслеживается генами через их настройку эпигенетикой.
Данную задачу до конца смогли решить и объяснить через исследование дрозофилы Drosophila melanogaster , она является одним из модельных организмов. Пошаговый контроль эмбриогенеза эксперименте с дрозофилой ( Эмбриогенез дрозофиллы ) визуализировали с помощью присоединения флуоресцентных красителей к каждому типу белка, который отвечал за тот или иной участок развития дрозофилы.
В 1994 году Вальтер Геринг обнаружил что ген pax 6, важный для формирования глаз у плодовых мух, в точности совпадает с геном формирования глаз у мышей и людей. Так же этот ген повторяется у кальмаров, головоногих моллюсков. Но это открытие противоречило прошлой теории о том, что глаза возникали 40 раз независимо друг от друга в живой природе и каждый из типов строения сильно отличается от других. Например глаз плодовой мухи состоит из сотен маленьких линзовидных структур. Человеческий глаз имеет слепое пятно там, где находится зрительный нерв, свет в итоге должен пройти через слой нервных волокон, прежде чем попасть на сетчатку. С этим связано то что человеческий глаз как бы перевёрнут. Напротив у головоногих глаза имеют сетчатку, затем слой нервных волокон и затем стенку глаза. Но все эксперименты с геномом показали что каждый раз у всех организмов именно ген pax 6 контролирует развитие глаз.
Это указывает на общего предка у всех организмов и является наглядным доказательством глубокой гомологии между структурами, до этого считавшихся конвергентно возникавшими и не связанными генетически.
Ограниченное количество эмбриональных генов
В организме всех животных имеющих двухстороннюю симметрию есть одинаковые, базовые гены контролирующие развитие от эмбриона до взрослого организма. Данные гены встречаются у всех животных и потому считаются одними из самых древних. У последнего общего предка Урбилатерия, было 7 Pax генов. они сохранены у всех последующих организмов и отвечают за те же функции развития организма, что и у предка.
| Урбилатерия? ↓ |
|
||||||||||||||||||

Различия в начале экспрессии разных генов Pax влияют на план строения тела, количество рук, ног и так далее. Эти гены отвечают за то, какими частями тела станут универсальные клетки - стволовые клетки. Эти клетки могут стать любыми частями тела, любыми тканями. Таким образом гены Pax отвечают за пространственную и временную симметрию организма, когда все части тела вырастают строго в нужных местах, начинают расти строго в нужно время и достигают окончания роста так же в нужное время.
При экспериментах было показано насколько сходство генов велико. Если взять ген Pax 6, который экспрессировался от дрозофилы и ввести его мыши, то у мыши так же вырастут глаза. Это наглядно доказывает что большая часть эволюции в морфологии животных основана не на появлении новых генов, а на изменений настроек этих генов = уровне метилирования, уровне экспрессии. Так же имеет место приобретения генами новых функций, при сохранении самих генов.

Потеря ног у змей и других животных хороший пример подобной регулировки генов. У змей не исчезал ген отвечающий на конечности, однако изменился радикально уровень его экспрессии, эпигенетика. Ген Distal-less очень слабо экспрессируется у змей или вообще не экспрессируется в тех областях, где формируются конечности у других теропод. Одновременно ген Distal-less определяет рисунок на крыльях бабочек Heliconius erato и Heliconius melpomene, что является примером изменений функции у данных генов. При этом бабочки по разному эволюционировали и узор стал получаться у них независимо друг от друга и ген который контролирует узор оба раза взял на себя эту функцию независимо.
Эти сохраняющиеся на протяжении сотен миллионов лет гены имеют тенденцию эволюционировать в одну и ту же функцию конвергентно. Гомеобоксные гены также встречаются у растений, подразумевая, что они являются общими для всех эукариот.
Регуляторные сети эмбрионов
Данный параметр напрямую связан с регуляцией экспрессии генов, в том числе с метилированием ДНК. В итоге белки регуляции используются повторно не через дублирование или модификацию, а посредством сложной настройки под конкретный процесс - плейотропию. Локусы таких белков-генов имеют сложные, крупные цис-регуляторные элементы. Например не имеющий плейотропии ген родопсина в плодовой мухе, имеет цис регуляторный элемент с несколькими сотнями пар оснований. Ген родопсина имеющий плейотропию содержит 6 цис регуляторных элементов и более 7 000 пар оснований. Подобный набор рождает целую сеть настроек. 67 транскрипционных факторов в плодовой мухе контролируют по 124 гена-мишени каждый. Такая сложная сеть позволяет контролировать развитие эмбриона с общей первичной клетки до полноценного организма. Гены включаются строго в нужное время в нужном месте. Часть генов отвечают за ферменты, ткани, органы эмбриона. Многие другие гены являются регуляторными, получается каскадно-замкнутая цепь реакции между белками, когда включение одного гена неизбежно заставляет включаться другие белки и процессы синхронизировано протекают во всех частях организма.
Именно на плодовой мухе получилось изучить столь сложную каскадно-регуляторную сеть. Этот же принцип, когда сеть из генов-белков настолько зависима от каждого элемента внутри себя, делает любые генетические мутации в ДНК вредными и естественный отбор убивает носителя таких мутаций. Пример подобной системы можно рассмотреть на белке бикоид, который тесно связан с рядом других белков при развитии, сегментировании тела плодовой мухи.
Bicoid стимулирует развитие переднего отдела эмбриона дрозофилы двумя путями. С одной стороны, он связывается с мРНК caudal, ингибируя её трансляцию и таким образом подавляя программу развития заднего отдела, которую активирует caudal. С другой стороны, он является транскрипционным фактором семейства Hox и связывается с регуляторными участками гена hunchback, активируя его. Для активации этого гена не нужны большие концентрации Bicoid, что также подтверждалось в экспериментах с нокаутом swallow и exuperantia: в таких мутантах нормально образовывались структуры торакса, но не головы. Было предсказано, что эти гены имеют энхансеры с меньшей аффинностью, что потом блестяще подтвердилось в эксперименте для генов buttonhead, empty spiracles и orthodenticle. Энхансеры этих генов также имеют сайт связывания Hunchback. Под регуляцией Bicoid также находится ген knirps, который экспрессируется из всей передней части зародыша только в акроне, поскольку одновременно его репрессирует hunchback. На этом примере видно, что именно разные пространственно-временные комбинации имеют решающее значение для тонких эффектов смены программы для отдельных клеток. Аналогично, ген evenskipped активируется Bicoid только во втором брюшном сегменте, так как там наблюдается требуемая низкая концентрация Bicoid, а также присутствует Hunchback.

Это обуславливает высокую стабильность генов отвечающих за сегментирование тела, его первичную настройку - положение органов, тканей. В итоге один и тот же ген в мышах и плодовой мухе отвечает за развитие конечностей.
Дальнейшее развитие ЭБР
В XXI веке стало ясно, что многообразие строения тел у разных животных не означает такое же разнообразие в последовательностях генов. Как отмечают Джон Герхарт и Марк Киршнер это является парадоксом. Учитывая важность Hox-генов они должны быть максимально разнообразны, но именно они в живых существах оказались наиболее консервативны и наименее подвержены изменениям. Получается что количество генов почти идентично, а их последовательность у мухи и лошади слишком мало отличается, что бы только эта последовательность объясняла столь разную структуру тела у животных. Так же Hox-гены не могут изменять строение тела через мутации, опять таки в силу своей консервативности. Объяснение идёт через механизм изменения настроек генов, без добавления новых генов, без изменения последовательности генов и называется - эпигенетический механизм.
Вариативность генов


Из вывода о важности эпигенетики, следует то что эволюция морфологии животных во многом идёт на уровне регуляции генов, а не на уровне добавления новых генов и изменения их последовательности. Один и тот же ген может быть отрегулирован по разному, например когда клюв большого наземного зяблика Дарвина увеличивается геном BMP или когда этот же ген отключает рост ног у змей. Так же ген может приобретать новую функцию и при этом отключается старая функция - один ген у разных животных контролирует либо рост нижней челюсти у позвоночных, либо ноги и усики у плодовой мухи, либо рисунок пятен на крыльях бабочки. В силу естественного отбора, если гены включались излишне сильно, то особь получала значительные изменения в структурах тела, а это всегда риск того что особь окажется не жизнеспособной и не сможет давать потомство. Таким образом любые неудачные варианты регуляции генов отсекаются естественным отбором. Так же в силу большого числа животных на планете срабатывает статистическая вероятность и получается конвергентная эволюция, когда разные виды бабочек Heliconius erato и Heliconius melpomene получают разную окраску крыльев, но ген отвечающий за это оба раза один и тот же. В итоге подобные изменения в одном и том же гене могут способствовать появлению новых видов.
Объединение эпигенетических изменений
Эволюционные инновации могут начинаться в ключе устаревшей теории Ламарка - Ламаркизме. Эти изменения на самом деле будут начинаться с эпигенетической регуляции генов, а в дальнейшем, в силу того что особь которая выжила, будет и дальше сохранять эпигенетические настройки генов которые позволили ей выживать, в итоге передаст настроенные гены потомкам и в дальнейшем настройки закрепляются уже на генном уровне, на уровне мутаций. Эпигенетика затрагивает модификацию ДНК путём обратимого метилирования. Метилирование в итоге это реакция каждого организма на условия окружающей среды, от температуры и влажности, до стрессов, голода и изменения высоты проживания, а значит атмосферного давления. Организмы изначально очень пластичны и способны подстраиваться под широкий спектр географических, физических и химических условий на Земле. По теории биологов Ньюмана и Мюллера именно данная особенность генов - их метилирование и изменение настроек в ответ на среду, позволяло выживать организмам на раннем этапе развития планет и эта же способность сохраняется и сегодня.
Подобная система согласуется с Расширенным эволюционным синтезом и в итоге даёт ряд прогнозируемых предсказаний для такой системы развития организмов:
1 - Изменение фенотипа может предшествовать изменению генотипа.
2 - Изменения в фенотипе преимущественно положительные ( в силу метилирования генов, в ответ на давление среды ), а не нейтральные и это не согласуется с с устаревшей ещё в 1990-х Нейтральной теорией молекулярной эволюции.
3 - Изменения фенотипа появляются во многих организмах, а не в одном организме.
4 - Революционное, ускоренное изменение фенотипа может происходить через мутацию, облегчённую вариацию ( использование ранее имевшихся генов ) или в силу пороговых событий - вымирание, катастрофы.
5 - Повторная эволюция в изолированных популяциях может идти путём конвергентной эволюции или смещения развития.
6 - Адаптация организма может происходить через естественный отбор, воздействие окружающей среды, не-генетическое наследование ( эпигенетическое ) и через культуру и обучение. Эффект Болдуина - это усвоенное поведение, ему обучают молодую особь и такой механизм начинает активно влиять на эволюцию вида.
7 - Быстрая эволюция может быть результатом одновременно воздействия окружающей среды, естественного отбора и общей динамики развития.
8 - Наследуемая изменчивость направлена на варианты, которые являются адаптивными и интегрированными с фенотипом.
9 - Устройство организмов в той или иной пищевой нише смещено в сторону изменения окружающей среды, в итоге конструкция лучше подходит под условия и под них же отбирается фенотип, что повышает общую приспособленность популяции.
10 - Родственный отбор - наиболее известные примеры это муравьи и пчёлы. При родственном отборе часть популяции жертвует своими силами и способностью к размножению в пользу особо успешных в размножении особей и тем самым вид успешно поддерживает свою численность.
11 - Многоуровневый отбор - это теория объясняющая крайне быстрое развитие человека по сравнению с животными. В теории идёт упор на то, что отбор в итоге подобен русской матрёшке. Сначала идут гены, затем клетки, организм и в конце - группы. В силу общего естественного отбора выживают те организмы и группы, в которых "матрёшка" многоуровневого отбора функционирует максимально согласованно, что увеличивает успех в размножении вида. Теория утверждает, что отбор и конкуренция между группами перевешивают конкуренцию отдельных организмов внутри каждой группы. Эта теория объясняет, почему вариации между группами людей больше, нежели вариации внутри групп людей. Эдвард Уилсон подвёл итог теории - Эгоистичные особи выигрывают у альтруистов внутри группы, но группа альтруистов победит группы, в которых много эгоистичных особей.
12 - Самоорганизация.
Генетическая ассимиляция
Адаптивное развитие
Это теория в составе ЭБР о том, что организм с самого начала своего зарождения реагирует на давление тех или иных факторов среды и изменяется в пределах вариативной изменчивости своего вида, так называемая норма реакции. В целом любой организм при развитии может пойти по пути генотипа своего родителя и это будет сохранение прошлой траектории развития с изменения в русле прошлых особенностей фенотипа - положительное развитие. Второй путь это отрицательный - в сторону от любых изменений в фенотипе и возможные изменения уже не в сторону прошлой линии. Либо организм идёт по абсолютному пути - изменения либо активно происходят, либо не происходят вообще.
Данная концепция сложно доказуема. Но например у брюхоногих моллюсков раковина по типу улитки всегда строится в виде трубки, но трубка может расти как в длину таки в диаметре. Причина этого в том что естественный отбор проходили разные типу улиток с разными параметрами раковин. В итоге выживали носители раков в форме спирали, каури, высокие башенные спирали.
Среди многоножек - сороконожки всегда имеют 15 сегментов составляющих тело во взрослом возрасте. Это указывает на смещение развития в сторону нечётного числа сегментов. Другой отряд - геофилы ( Geophilomorpha ) имеют большее число сегментов тела, от 27 до 191, но число всегда нечётное, в итоге все нечётные чсила в этом диапазоне заняты тем или иным видом.
Эво-дево и концепция опухолей в эволюции
Это дальнейшее осмысливание и исследования на тему теории эво-дево. Основная работа ведётся под руководством профессора А.П. Козлова. Доработанная теория эво дево не противоречит биологическим теориям которые общепризнаны. В том числе эво дево не вступает в конфликт ни с дарвинизмом ни с синтетической теорией эволюции. Она просто заполняет лакуны между разными направлениями биологии и решает ряд пока не объяснённых научно задач. Основная идея - опухоли это почти всегда не смертельные для носителя образования и они могут принимать участие в возникновении новых типов клеток, тканей, органов.
Многоклеточные организмы в процессе эволюции и просто развития нуждаются в непрерывном источнике дополнительных клеточных масс. Такие клетки должны обладать высокими биосинтетическими и морфологическими потенциалами, только тогда возможна прогрессивная эволюция тканей, особенно это критично для хордовых. Однако лишних клеток при стандартном онтогенезе не образовывается просто в силу настроек ДНК и эпигенетики. Наличие универсальных стволовых клеток не решает этот вопрос, они уже встроены в онтогенез и регулируются функциональными петлями обратной связи.
Опухоли имеют ряд важный параметров. Главный - есть виды опухолей точно наследуемых, это главный фактор для внесения вклада в эволюцию, поскольку эволюция одного организма без возможности передачи генов не имеет смысла в эволюционной биологии. Опухоли наследственного типа могут представлять собой относительно неустойчивые переходные формы, они связывают различные уровни сложности эволюции клеток. Их стабилизация в дальнейшем была достигнута через экспрессию новых генов и комбинаций генов.
Фенотип при развитии от зиготы и до окончания роста крайне чувствителен к любым изменениям в генетике и морфологии, они могут привести к смерти организма. Опухоли решают вопрос гиперметилированием внутри своих клеток. Так же опухоль это не единый организм-паразит, а конкурирующие между собой слои клеток. Это и осложняет лечение онкологических заболеваний, клетки любой опухоли не точная копия друг друга, а скорее соперники в борьбе за ресурсы организма. Это же закладывает вероятность положительного действия опухолей в разных эволюционных задачах.
См. также
Литература
На русском языке
- Рэфф Р., Кофмен Т. Эмбрионы, гены и эволюция: Пер. с англ. — М.: Мир, 1986. — 404 с.
На английском языке
- Buss L. W. The Evolution of Individuality. — Princeton University Press. — ISBN 978-0-691-08468-8.
- Carroll Sean B. Endless Forms Most Beautiful: The New Science of Evo Devo and the Making of the Animal Kingdom. — Norton, 2005. — ISBN 978-0-393-06016-4.
- Goodwin B. How the Leopard Changed its Spots. — 1994. — ISBN 978-0-691-08809-9.
- Keywords and Concepts in Evolutionary Developmental Biology. — New Delhi, India: Discovery Publishing House, 2007. — ISBN 978-81-8356-256-0.
- Kirschner M., Gerhart J. The Plausibility of Life: Resolving Darwin's Dilemma. — Yale University Press, 2005. — ISBN 978-0-300-10865-1.
- From Embryology to Evo-Devo: A History of Developmental Evolution / Laubichler M. D., Maienschein J., ed. — The MIT Press, 2007. — ISBN 978-0-262-12283-2.
- Minelli A. The Development of Animal Form: Ontogeny, Morphology, and Evolution. — Cambridge University Press, 2003. — ISBN 978-0-521-80851-4.
- Orr H. A. Turned on: A revolution in the field of evolution? // The New Yorker. — 2005. Discussion of Carroll, Endless Forms Most Beautiful
- Raff R. A. The Shape of Life: Genes, Development, and the Evolution of Animal Form. — The University of Chicago Press, 1996. — ISBN 978-0-226-70266-7.
- Sommer, R. J. The future of evo–devo: model systems and evolutionary theory // Nature Reviews Genetics. — 2009. — Т. 10, вып. 6. — С. 416–422. — doi:10.1038/nrg2567. — PMID 19369972.
Ссылки
- Scott F. Gilbert, The morphogenesis of evolutionary developmental biology
- Tardigrades (water bears) as evo-devo models, a short video from NPR’s Science Friday
- Блог — учебник по Evo-devo, [1]
| Эволюционные процессы | |
|---|---|
| Факторы эволюции | |
| Генетика популяций | |
| Происхождение жизни | |
| Исторические концепции | |
| Современные теории | |
| Эволюция таксонов | |