
| Цитохром-c-оксидаза | |
|---|---|
 Цитохром с-оксидаза быка. | |
| Идентификаторы | |
| Шифр КФ | 7.1.1.9 |
| Номер CAS | 9001-16-5 |
| Базы ферментов | |
| IntEnz | IntEnz view |
| BRENDA | BRENDA entry |
| ExPASy | NiceZyme view |
| MetaCyc | metabolic pathway |
| KEGG | KEGG entry |
| PRIAM | profile |
| PDB structures | RCSB PDB PDBe PDBj PDBsum |
| Gene Ontology | AmiGO • EGO |
| Поиск | |
| PMC | статьи |
| PubMed | статьи |
| NCBI | NCBI proteins |
| CAS | 9001-16-5 |
Цитохром-c-оксида́за (цитохромоксидаза) или цитохром-c-кислород:оксидоредуктаза, также известная как цитохром aa3 и комплекс IV — терминальная оксидаза аэробной дыхательной цепи переноса электронов, которая катализирует перенос электронов с цитохрома с на кислород с образованием воды. Цитохромоксидаза присутствует во внутренней мембране митохондрий всех эукариот, где её принято называть комплекс IV, а также в клеточной мембране многих аэробных бактерий.
Комплекс IV последовательно окисляет четыре молекулы цитохрома с и, принимая четыре электрона, восстанавливает O2 до H2O. При восстановлении O2 четыре H+ захватываются из митохондриального матрикса для образования двух молекул H2O, а ещё четыре H+ активно перекачиваются через мембрану. Таким образом, цитохромоксидаза вносит свой вклад в создание протонного градиента для синтеза АТФ и является частью пути окислительного фосфорилирования. Кроме того, этот мультибелковый комплекс играет ключевую роль в регуляции активности всей дыхательной цепи и производстве энергии эукариотической клеткой.
История изучения
Цитохромоксидаза была открыта ирландским врачом и учёным Ч. А. МакМанном, который в 1885 году описал обратимые изменения в спектре поглощения при длине волны в 605 нм, происходящие при окислении в клетках животных, что является характерной спектральной подписью цитохромоксидазы. Однако его работы были раскритикованы влиятельными физиологами Гоппе-Зейлером и Леви, которые постулировали, что МакМанн просто наблюдал поглощение продуктов распада гемоглобина. В результате исследования этого фермента прекратились более чем на 30 лет, пока в 1923 году Ханс Фишер не подтвердил результаты МакМанна.
Дальнейшие исследования этого фермента были продолжены немецким учёным Отто Варбургом. В своей работе он ингибировал дыхание в суспензии дрожжей при помощи CO, а затем получал спектры поглощения, снимая ингибирование путём облучения когерентным пучком света с разной длиной волны. Из полученных данных следовало, что ингибируемый фермент — гемопротеин, в котором гем находится в комплексе с CO. Варбург связал новый, неизвестный белок с функцией клеточного дыхания и применил к нему используемый им с 1924 года термин Atmungsferment или «дыхательный фермент». Работа была опубликована в 1929, а в 1931 Варбург получил за неё Нобелевскую премию по физиологии и медицине с формулировкой «за открытие природы и механизма действия дыхательного фермента».
Немалый вклад в понимание природы комплекса IV внёс британский учёный Дэвид Кейлин. В 1939 году, в совместной работе с Е. Ф. Хартри он обнаружил неизвестный ранее цитохром, получивший название a3, который обладал способностью окислять цитохром c. Новый цитохром обладал тем же спектром поглощения, что и загадочный дыхательный фермент Варбурга, а также ингибировался под действием СО и KCN. В своей работе Кейлин ввёл в обиход название цитохром c-оксидаза, предложенное Малкольмом Диксоном в 1928 году. Варбург и Кейлин длительное время спорили о природе циохромоксидазы: Варбург считал, что кофактором этого фермента может быть только железо, в то время как Кейлин полагал, что это медьсодержащий белок. По прошествии лет оказалось, что оба великих учёных были правы: в состав цитохромоксидазы входит как железосодержащий гем, так и атом меди.
Механизм связывания цитохромоксидазой кислорода исследовал американский биохимик Бриттон Ченс, который в середине 70-х годов двадцатого века, используя передовую технику ЯМР и спектроскопии при низких температурах, обнаружил фермент-субстратный комплекс цитохромоксидазы — аддукт гема a3 с молекулярным кислородом.
В 1977 году финский учёный Мартин Викстрём показал, что цитохромоксидаза в процессе своей работы перекачивает протоны через мембрану, чего долгое время не мог принять создатель хемиосмотической гипотезы, Питер Митчелл. Тем не менее накапливавшиеся экспериментальные данные свидетельствовали в пользу правоты Викстрёма, и позже Митчел признал свою ошибку.
Первые попытки выделить фермент предпринимались начиная с 1941 года: поскольку тогда ещё не было разработано процедур для выделения больших мембранных белков, то приходилось действовать путём проб и ошибок. В первых процедурах выделения использовались соли желчных кислот, что вызывало большие потери активности. Появление неионных детергентов наподобие Triton X-100 вызвало новый подъём в этой области с 1966 по 1974 год и позволило получить первые чистые препараты. Первая трёхмерная структура с атомным разрешением комплекса появилась чуть позже, в 1995 году.
Структурная организация комплекса IV
Комплекс IV из митохондрий млекопитающих и птиц состоит из 13 белковых субъединиц, три из которых обладают каталитической активностью, связывают кофакторы и кодируются генами митохондрий (исключение составляет субъединица III у Chlamydomonas reinhardtii и Polytomella sp, которая кодируется в ядре). Остальные десять субъединиц закодированы в ДНК ядра. В 2012 году поступило сообщение об обнаружении 14-й субъединицы, но позже оно было опровергнуто. В мембране митохондрий комплекс существует в виде гомодимера, каждый мономер состоит из 13 субъединиц. Молекулярная масса такого димера, выделенного из митохондрий быка, составляет приблизительно 350 кДа. Встречающиеся в мембране немногочисленные мономеры обладают вдвое более высокой каталитической активностью.
У S. cerevisiae комплекс IV состоит всего из 11 субъединиц, однако отсутствующие субъединицы в бычьем комплексе представляют собой маленькие периферические белки, так что дрожжевая цитохромоксидаза существенно не отличается от таковой у млекопитающих. Значительно меньше известно о комплексе IV у растений, он и по сей день остаётся одним из самых неизученных комплексов растительных митохондрий. Последние эксперименты по его выделению из арабидопсиса и исследование его методом голубого нативного электрофореза показали, что он, по-видимому, состоит из восьми субъединиц, схожих с субъединицами комплекса IV других эукариот, и шести дополнительных субъединиц, специфичных для растений. Менее точное разделение комплекса IV из картофеля и фасоли дало рисунок полос, схожий с таковым у арабидопсиса: можно точно сказать, что их комплекс IV состоит по крайней мере из 9—10 субъединиц. Бактериальные комплексы существуют в мембране в виде мономеров и состоят из 3—4 субъединиц, три из которых гомологичны трём закодированным в митохондриях субъединицам эукариот.
Субъединицы

Три большие субъединицы комплекса (I—III), гомологичные бактериальным, несут на себе все необходимые кофакторы и осуществляют основные реакции катализа, связанные, в том числе, и с переносом протонов. Расположенные на периферии малые ядерные субъединицы не участвуют в этом процессе. В настоящее время специфические функции известны только для четырёх ядерных субъединиц (IV, Va, VIa-L, VIa-H), но очевидно, что все они играют роль в сборке, димеризации, а также регуляции активности комплекса. Ядро комплекса IV обладает крайне высокой каталитической активностью, которая подавляется плотно связанными с ним вспомогательными ядерными субъединицами, что особенно важно для регуляции всего дыхания в целом. У позвоночных многие из этих субъединиц представлены несколькими тканеспецифичными изоформами, каждая из которых кодируется отдельным геном. Экспрессия каждой изоформы зависит от типа ткани, стадии развития организма и может изменятся в зависимости от внешних условий, что позволяет чётко регулировать снабжение энергией разных органов и тканей.
Появление большого разнообразия ядерных субъединиц после полногеномной дупликации у позвоночных приблизительно совпадает по времени с утратой ими альтернативной оксидазы, которая обеспечивала альтернативный путь для электронов к кислороду в обход комплекса IV. Роль этих субъединиц особенно возросла поскольку клетки млекопитающих утратили способность переключаться между разными терминальными оксидазами, как это происходит у прокариот. Например, E. coli имеет две терминальные хинон-оксидазы; при нормальном содержании кислорода она преимущественно экспрессирует цитохром bo3, а при низком переходит на цитохром bd, который обладает повышенным сродством к кислороду, но не перекачивает протоны. Очевидно, что в таких условиях ядерные субъединицы приняли на себя функцию по управлению активностью всего окислительного фосфорилирования в зависимости от уровня кислорода.
Субъединица Va специфически связывает тиреоидный гормон 3,5-дийодотиронин, но не взаимодействует с тироксином или трийодтиронином. В результате такого взаимодействия комплекс IV перестаёт аллостерически ингибироваться АТФ. Этот механизм объясняет кратковременный стимулирующий эффект тиреоидных гормонов на метаболизм млекопитающих.
У млекопитающих субъединица IV-2 экспрессируется в основном в мозге и лёгких, а в остальных тканях её синтез индуцируется в условиях гипоксии. У рыб эта изоформа сильнее экспрессируется в жабрах. Хотя у всех позвоночных есть по одной копии обеих изоформ субъединицы IV, активация экспрессии IV-2 в ответ на недостаток кислорода есть только у млекопитающих и отсутствует у рыб и рептилий, а у птиц ген COX4-2, кодирующий изоформу IV-2, не функционален. У мышей, нокаутных по гену IV-2, наблюдались трудности в сокращении дыхательных путей, пониженное содержание АТФ в лёгких, а с возрастом появлялись патологии дыхательной системы, включая кристаллы Шарко-Лейдена. Эти экспериментальные данные свидетельствуют о важности изоформы IV-2 для нормальной работы лёгких млекопитающих.
Для субъединиц VIa-L и VIa-H удалось определить специфические функции. Оказалось, что способность перекачивать протоны (стехиометрия H+/e−) у комплекса из почек и печени снижалась с 1 до 0,5 низкими концентрациями свободной пальмитиновой кислоты, чего не происходило с комплексом IV из сердца и мышц, содержащего изоформу VIa-H. Предположительное физиологическое значение этого процесса заключается в усилении термогенеза и поддержании температуры тела во всех тканях кроме мышечной в ответ на свободный пальмитат. Субъединица VIa-H из сердца и мышц стимулирует работу комплекса, связывая АДФ, и наоборот, снижает стехиометрию H+/e− при высоком соотношении АТФ/АДФ. Физиологическое значение этой особенности заключается в усилении термогенеза в мышцах во время сна или отдыха, когда расход АТФ снижен, а соотношение АТФ/АДФ остаётся высоким. Субъединица VIa-H отсутствует у рыб.
| Субъединица | Изоформа | Белок | Описание |
|---|---|---|---|
| I | - | Cox1 | Связывает гем а, гем а3, центр CuB, имеет протонные каналы. |
| II | - | Cox2 | Связывает центр CuA, взаимодействует с цитохромом с. |
| III | - | Cox3 | Стабилизирует транспорт протонов. |
| IV | IV-1 IV-2 |
Cox41 | Обеспечивает аллостерическое ингибирование АТФ. |
| Cox42 | Экспрессируется в основном в лёгких, плаценте и мозге, индуцируется гипоксией. Возможно, O2-зависимое ингибирование АТФ. | ||
| Va | - | Cox5a | Связывает 3,5-дийодотиронин, в результате чего снимается ингибирование АТФ. |
| Vb | - | Cox5b | Связывает Zn2+. |
| VIa | VIa-L VIa-H |
Cox6a1 | Печёночая изоформа. Экспрессируется во всех тканях, кроме скелетных мышц и сердца. Снижает стехиометрию H+/e− с 1 до 0,5 в присутствии пальмитата. |
| Cox6a2 | Сердечная изоформа. Экспрессируется в сердце и скелетных мышцах. Снижает стехиометрию H+/e− с 1 до 0,5 при высоком соотношении АТФ/АДФ. | ||
| VIb | VIb-1 VIb-2 |
Cox6b1 | Во всех тканях. Обеспечивает димеризацию комплекса. |
| Cox6b2 | Специфична для семенников. Возможно увеличивает интенсивность дыхания. | ||
| VIc | - | Cox6c | Во всех тканях. |
| VIIa | VIIa-L VIIa-H VIIa-R SIG81 |
Cox7a2 | Экспрессируется во всех тканях, кроме скелетных мышц и сердца. |
| Cox7a1 | Экспрессируется в сердце и скелетных мышцах. | ||
| Cox7a3 | - | ||
| Cox7A2L | - | ||
| VIIb | VIIb-1 VIIb-2 |
Cox7b | Во всех тканях. |
| Cox7b2 | Специфична для семенников. Возможно увеличивает интенсивность дыхания. | ||
| VIIc | - | Cox7c | Во всех тканях. |
| VIII | VIII-L VIII-H VIII-3 |
Cox8a | Во всех тканях. |
| Cox8b | Экспрессируется в скелетных мышцах и буром жире. У человека превратилась в псевдоген. | ||
| Cox8c | - |
Кофакторы
Кофакторы комплекса IV расположены на двух крупных единицах I и II, встроенных в мембрану. Субъединица I образует двенадцать трансмембранных α-спиралей и содержит три окислительно-восстановительных центра: гем а (окислительно-восстановительный потенциал + 0,22 В) и так называемый биядерный центр a3-CuB, в состав которого входит гем а3 и атом меди CuB. Химически гемы a и a3 идентичны, но железо гема а шестикоординированно, поскольку образует шесть координационных связей с четырьмя атомами азота пиррольных колец и двумя атомами азота близлежащих остатков гистидина, а в геме а3 оно образует только пять координационных связей, что делает шестую связь доступной для связывания с молекулярным кислородом. Напротив железа гема а3 располагается атом меди CuB, лигированный тремя остатками гистидина. Хотя между железом и медью биядерного центра нет связующих элементов, между ними наблюдается сильное антиферромагнитное сопряжение. Окислительно-восстановительный потенциал биядерного центра составляет приблизительно + 0,24 В.
Кристаллографические исследования выявили необычную посттрансляционную модификацию субъединицы I: гистидин-240ковалентно связан через свой атом азота в тау-положении с мета-углеродом бензольного кольца тирозина-244. Этот остаток тирозина поставляет электрон и протон для восстановления кислорода с образованием нейтрального радикала. Кроме того, ковалентная связь создаёт пентамерное кольцо из аминокислот, остаток глутамата которого является важным компонентом транспорта протонов.
На субъединице II расположен CuA-центр (редокс-потенциал = − 0,70 В), который состоит из двух атомов меди, напрямую соединённых ковалентной связью. Он лигирован шестью остатками аминокислот: двумя остатками цистеина, двумя остатками гистидина, одним остатком метионина и пептидным карбоксилом глутаминовой кислоты. Функционирует как одноэлектронный переносчик.
Методами рентгеноструктурного анализа и сайтспецифичного мутагенеза субъединицы I были выявлены пути, по которым протоны могут проникать через комплекс и пересекать мембрану. Эти пути получили название D-, K- и Н-каналов. В каналах, выстланных полярными аминокислотными остатками, удерживается разное число молекул воды. Обнаруженный в комплексе ион Mg2+, возможно, как раз и нужен для стабилизации этих молекул. Предполагается, что К-канал связывает водную фазу матрикса с биядерным центром и служит для доставки «субстратных» протонов, необходимых для образования воды из кислорода. D-канал, по-видимому, формирует сквозной путь, и по нему могут проходить как «субстратные» протоны, так и протоны, перекачиваемые через мембрану. У эукариот обнаружен дополнительный Н-канал, который, вероятно, также является сквозным.
Реакция
Суммарная реакция, катализируемая комплексом, описывается следующим уравнением:
- 4цит. c2+ + O2 + 8H+in → 4цит. c3+ + 2H2O + 4H+out
Путь электрона в комплексе известен. Цитохром с связывается на субъединице II при посредничестве субъединиц I, III и VIb и восстанавливает CuA-центр, расположенный вблизи поверхности мембраны. С CuA-центра электрон уходит на гем а и далее на биядерный центр a3-CuB, расположенные в толще мембраны. Именно в биядерном центре происходит связывание О2 и его восстановление до Н2О. Поскольку кислород обладает высоким сродством к электронам, то в процессе восстановления до воды он высвобождает большое количество свободной энергии. Благодаря этому аэробные организмы способны получать гораздо большее количество энергии, чем можно выработать исключительно анаэробным способом.
Механизм восстановления кислорода
Механизм восстановления кислорода уже давно является предметом интенсивного изучения, но ясен не до конца. Каталитический цикл цитохромоксидазы состоит из шести стадий, обозначаемых A (аддукт, англ. Adduct), P (пероксиинтермедиат от англ. Peroxy intermediate), F (феррилоксоинтермедиат от англ. Ferryl-oxo intermediate), OH (полностью окисленное высокоэнергетическое состояние от англ. Fully-Oxidized High-Energy state), E (одноэлектронно-восстановленное состояние от англ. One-electron reduced state) и R (восстановленное состояние от англ. Reduced state) и названных так по состоянию биядерного центра. Следует отметить, что номенклатура каталитических состояний значительно устарела, не всегда отражает реальное химическое состояние биядерного центра и сохраняется во многом по историческим причинам. Так например, на стадии P кислород в биядерном центре находится совсем не в пероксидой форме, как то полагали 30 лет назад, а в оксоферрильном состоянии, где связь между атомами кислорода уже разорвана. Согласно современным представлениям, восстановление кислорода в цитохром-с-оксидазе происходит путём быстрого и полного восстановления с попарным переносом электронов, что исключает образование активных форм кислорода. Происходит следующая последовательность событий:
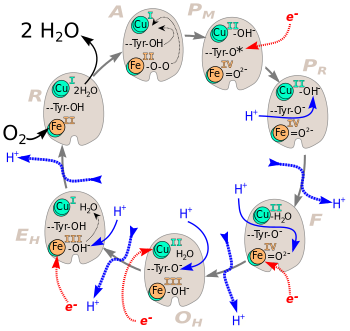
- A Полностью восстановленный биядерный центр быстро связывает O2 c образование кислородного аддукта, что приводит к конформационным перестройкам (обозначены тонкими чёрными стрелочками).
- PM Происходит быстрый перенос четырёх электронов на кислород: два поставляются железом гема а3 (FeII→FeIV), ещё один расположенным рядом CuB (CuI→CuII), а четвёртый приходит от остатка тирозина-244, он же отдаёт протон, необходимый для разрыва двойной связи O2. Образовавшийся нейтральный тирозиновый радикал восстанавливается до состояния аниона за счёт электрона от цитохрома с.
- PR Происходит протонирование Cu(II)-OH− с образованием молекулы воды.
- F Образовавшаяся молекула воды связывается с CuB координационной связью. Железо Fe(IV)=О2- восстанавливается до FeIII, а связанный с ним кислород протонируется. Высвобождается первая молекула воды.
- OH Тирозиновый анион протонируется, а CuB восстанавливается до CuI за счёт электрона от цитохрома с.
- EH Железо восстанавливается до FeII, после чего связанная с ним OH- группа протонируется с образованием второй молекулы воды.
- R В этом состоянии биядерный центр полностью восстановлен и комплекс готов к связыванию новой молекулы кислорода.
Механизм транспорта протонов
Известно, что эукариотическая цитохромоксидаза переносит через мембрану по одному протону на каждый электрон, полученный от цитохрома с. За один раз комплекс закачивает один «субстратный» протон, используемый для образования воды, через канал К и переносит один дополнительный протон через мембрану по каналу D. В ходе одного каталитического цикла акт транслокации приходятся на четыре относительно стабильных стадии: PM, F, OH, и EH.

Точный механизм транспорта протонов до сих пор остаётся не ясным: за последние годы было предложено множество моделей, в которых предпринимались попытки детально описания этого процесса. Не понятно и то, каким образом осуществляется сопряжение энергии электрона с перемещением протонов. Тем не менее, в общем виде это можно описать следующим образом:
- В начальной стадии цикла протонные каналы комплекса закрыты, затем цитохром с передаёт электрон на CuA-центр.
- Электрон быстро перемещается c CuA-центра на гем a, что ведёт к изменению окислительно-восстановительного потенциала и заставляет молекулы воды в канале D переориентироваться, делая его открытым для протона. В результате перемещения электрона с CuA на гем a происходит перемещение протона через канал D и его загрузка в сайт загрузки протона PLS (англ. proton loading site).
- Электрон переходит на биядерный центр к гему a3, в результате чего через канал K входит один субстратный протон. При этом протон в PLS испытывает значительное увеличение его кислотности (с pK=11 до pK=5).
- На завершающей стадии цикла предзагруженный в PLS протон выбрасывается наружу, как полагают, по причине электростатического отталкивания от субстратного протона, который участвует в восстановлении кислорода в биядерном центре.
Регуляция и сборка
Биогенез комплекса IV — очень сложный и хорошо регулируемый процесс, который уже длительное время является предметом усиленного изучения. В сборке комплекса участвует более двадцати закодированных в ядре вспомогательных факторов, а также белков, которые вставляют в него гемы а, а3 и атомы меди. Сюда же относятся по крайней мере 15 белков-активаторов трансляции митохондриальных субъединиц, ответственных за правильную транскрипцию и сплайсинг мРНК и активацию трансляции, специальные транслоказы, необходимые для транспорта ядерных субъединиц в митохондрии, а также ферменты биосинтеза кофакторов. Помимо специальных факторов сборки для биогенеза комплекса IV требуется ещё немалое количество белков с большой специфичностью, включая АТФ-зависимые пептидазы, ответственные за процессинг пропептидов.
Посттрансляционная регуляция активности комплекса IV не менее сложна и достигается за счёт множества разных способов. Сюда относятся фосфорилирование субъединиц, обратимое связывания части периферических субъединиц, регуляция за счёт использования определённых изоформ ядерных субъединиц, которое зависит от стадии развития и типа ткани, аллостерическое регулирование за счёт АТФ и АДФ в десяти сайтах связывания (у цитохромоксидазы млекопитающих), моно- и димеризация комплекса, а также его взаимодействие с другими дыхательными комплексами с образованием респирасом.
Фосфорилирование субъединиц комплекса имеет особую важность, поскольку связывает его активность с действием регуляторных каскадов клетки и работой цикла Кребса. Фосфорилирование и дефосфорилирование вызывает такие эффекты, как снятие ингибирования через АТФ во время стресса или запуск апоптоза. Всего в комплексе обнаружено 18 позиций для фосфорилирования, но точная функция фосфорилирования по каждому из этих положений не определена.
Положение в системе классификации белков
Цитохромоксидаза относится к белковому суперсемейству гем-медных оксидоредуктаз (в классификации ферментов перенесена в 7 класс - транслоказы), в которое входит большинство известных на данный момент терминальных оксидаз, а также редуктазы оксида азота(II), которые катализирую двухэлектронное восстановление NO до N2O с образованием воды. Для всех представителей этого суперсемейства характерно наличие субъединицы I с консервативной третичной структурой, одного низкоспинового гема и биядерного центра из атома меди и высокоспинового гема. Члены суперсемейства подразделяются на семейства по типу гема, наличию дополнительных кофакторов, аминокислотной последовательности, третичной структуры и количества субъединиц, типу окисляемого субстрата, а также по строению протон-переносящих каналов или отсутствию таковых. Наличие дополнительных субъединиц, несущих добавочные гемы или атомы металлов (или же полное отсутствие таковых) даёт возможность этим ферментам получать электроны от разного типа субстратов: разнообразных мембранных переносчиков типа хинонов, водорастворимых цитохромов или голубых медьсвязывающих белков.
Семейство А — самое большое и самое изученное семейство из всех гем-медных оксидоредуктаз. Для него характерны состав гемов типа aa3 или caa3. Представители этого семейства обычно состоят из трёх субъединиц: I, II и III, которые гомологичны субъединицам типового представителя семейства — митохондриальной цитохром-с-оксидазе. Они обладают как минимум двумя протонными каналами, D и K, и транслоцируют протоны со стехиометрией H+/e-. Цитохром-с-оксидаза млекопитающих относится к подсемейству А1 вместе с цитохромоксидазами P. denitrificans и R. sphaeroides.
Оксидазы из семейства В состоят из трёх субъединиц: I, II и IIa. Субъединица IIa представляет собой единственную трансмембранную цепь, аналогичной по структуре второй трансмембранной цепи субъединицы II из семейства А. Имеют только один альтернативный протонный канал К, стехиометрия переноса протонов — 0,5-0,75 H+/e-. Характерен набор гемов типа ba3, b(o)a3 и aa3.
К семейству C относятся только терминальные оксидазы типа cbb3. Обладают дополнительной субъединицей, которая может связывать один или два гема c. Это второе по размеру семейство кислород-редуктаз (24 %) после семейства А (71 %). Имеется альтернативный канал К, который отличается по строению от К-канала редуктаз из семейства B. Стехиометрия переноса протонов составляет 0,2-0,4 H+/e-, но по другим данным 0,6-1. Это семейство встречается только среди бактерий, так как большинство архей не умеют синтезировать гем с.
На основе биоинформатического анализа было предложено выделить малые семейства D, E, F, G, и H, которые представлены только у архей и обладают чрезвычайным разнообразием. В классической системе все эти семейства включены в состав семейства B, однако высокое разнообразие их первичной структуры говорит в пользу выделения их в отдельные семейства.
Внутриклеточное распределение
Три коровые субъединицы цитохром-с-оксидазы, закодированные в митохондриальном геноме, недавно были обнаружены за пределами митохондрий. Их нашли в зимогенных гранулах ацинусов поджелудочной железы. Относительно высокую концентрацию этих субъединиц обнаружили в секреторных гранулах вместе с гормоном роста в передней доле гипофиза. Функции этих субъединиц за пределами митохондрий ещё не определены. Кроме субъединиц цитохром-с-оксидазы вне митохондрий были обнаружены многие другие митохондриальные белки. В связи с этими находками была высказана гипотеза о существовании неизвестного механизма транспорта белков из митохондрий в другие клеточные компартменты.
Ингибиторы
Цианиды, сульфиды, азиды, монооксид углерода и монооксид азота связываются с окисленным или восстановленным биядерным центром фермента и конкурируют с кислородом, ингибируя при этом фермент, что приводит к смерти клеток от химической асфиксии. Метанол, который входит в состав технического спирта, в организме преобразуется в муравьиную кислоту, которая тоже может ингибировать цитохромоксидазу.
Клиническое и практическое значение
Мутации, затрагивающие ферментативную активность или структуру цитохром-с-оксидазы, приводят к тяжёлым и, как правило, фатальным нарушениям метаболизма. Такие нарушения обычно проявляются в раннем детстве и влияют преимущественно на ткани с высоким потреблением энергии (мозг, сердце, мышцы). Среди множества митохондриальных заболеваний, заболевания, связанные с дисфункцией или нарушением сборки цитохромоксидазы, считаются самыми тяжёлыми.
Подавляющее большинство нарушений работы цитохромоксидазы связаны с мутациями закодированных в ядре факторах сборки этого комплекса. Они обеспечивают правильную сборку и работу комплекса и участвуют в нескольких жизненно важных процессах, включая транскрипцию и трансляцию митохондриальных субъединиц, процессинг пропептидов и их встраивание в мембрану, а также биосинтез кофакторов и закрепление их в комплексе.
По состоянию на текущий момент удалось идентифицировать мутации в семи факторах сборки: SURF1, SCO1, SCO2, COX10, COX15, COX20, COA5 и LRPPRC. Мутации в этих белках могут приводить к изменению работы комплекса, неправильной сборке субкомплексов, нарушению транспорта меди или регуляции трансляции. Мутация в каждом из генов связана с этиологией определённого заболевания, некоторые из которых могут переходить в множественные расстройства. К такого рода генетическим нарушениям относятся синдром Лея, кардиомиопатия, энцефалопатия, лейкодистрофия, анемия и нейросенсорная тугоухость.
Гистохимия
Гистохимическое окрашивание комплекса IV используется для картирования метаболически активных участков мозга животных, поскольку существует прямая зависимость между активностью этого фермента и активностью всего нейрона. Такое картирование проводилось на мутантных мышах с различными нарушениями работы мозжечка, в частности на мышах из линии reeler и на трансгенной модели болезни Альцгеймера. Эта техника также применяется для картирования областей мозга животных, активных в процессе обучения.
ДНК-баркодирование
Последовательность участка гена субъединицы I цитохром-с-оксидазы (длиной порядка 600 нуклеотидов) широко используется в проектах, связанных с ДНК-баркодированием — определением принадлежности организма к тому или иному таксону на основе коротких маркеров в его ДНК.
См. также
- НАДН-дегидрогеназный комплекс
- Сукцинатдегидрогеназа
- Цитохром-b6f-комплекс
- Цитохром-bc1-комплекс
- Респирасомы
Источники
Литература
- Физиология растений / Под ред. И. П. Ермакова. — М.: Академия, 2005. — 634 с.
- Дэвид Л. Нельсон, Майкл М. Кокс. Основы биохимии Ленинджера. Биоэнергетика и метаболизм. = Leninger Principles of Biochemistry. — Бином. Лаборатория знаний, 2012. — Т. 2. — С. 344. — 692 с. — (Лучший зарубежный учебник). — ISBN 978-5-94774-365-4.
Ссылки
- Instituto Gulbenkian de Ciência & Universidade Nova de Lisboa. Heme-copper oxygen reductases (HCOs) classifier (сентябрь 2010).
- UMich Orientation of Proteins in Membranes families/superfamily-4
- MeSH Cytochrome-c+Oxidase